| Research article |
|


|
|
|
| Branch architecture of Tetraena mongolica Maxim. controls particle size distribution of nebkha sediments |
ZHAI Bo1, DANG Xiaohong2,*( ), LIU Jing3, LIU Xiangjie4, CHEN Xiaona4, LIU Yajing4 ), LIU Jing3, LIU Xiangjie4, CHEN Xiaona4, LIU Yajing4 |
1 School of Geographical Sciences and Planning, Jining Normal University, Ulanqab 012000, China
2 College of Desert Science and Engineering, Inner Mongolia Agricultural University, Hohhot 010018, China
3 Institute of Water Resources for Pastoral Area of the Ministry of Water Resources of China, Hohhot 010020, China
4 Experimental Center for Desert Forestry, Chinese Academy of Forestry, Bayannur 015200, China |
|
|
|
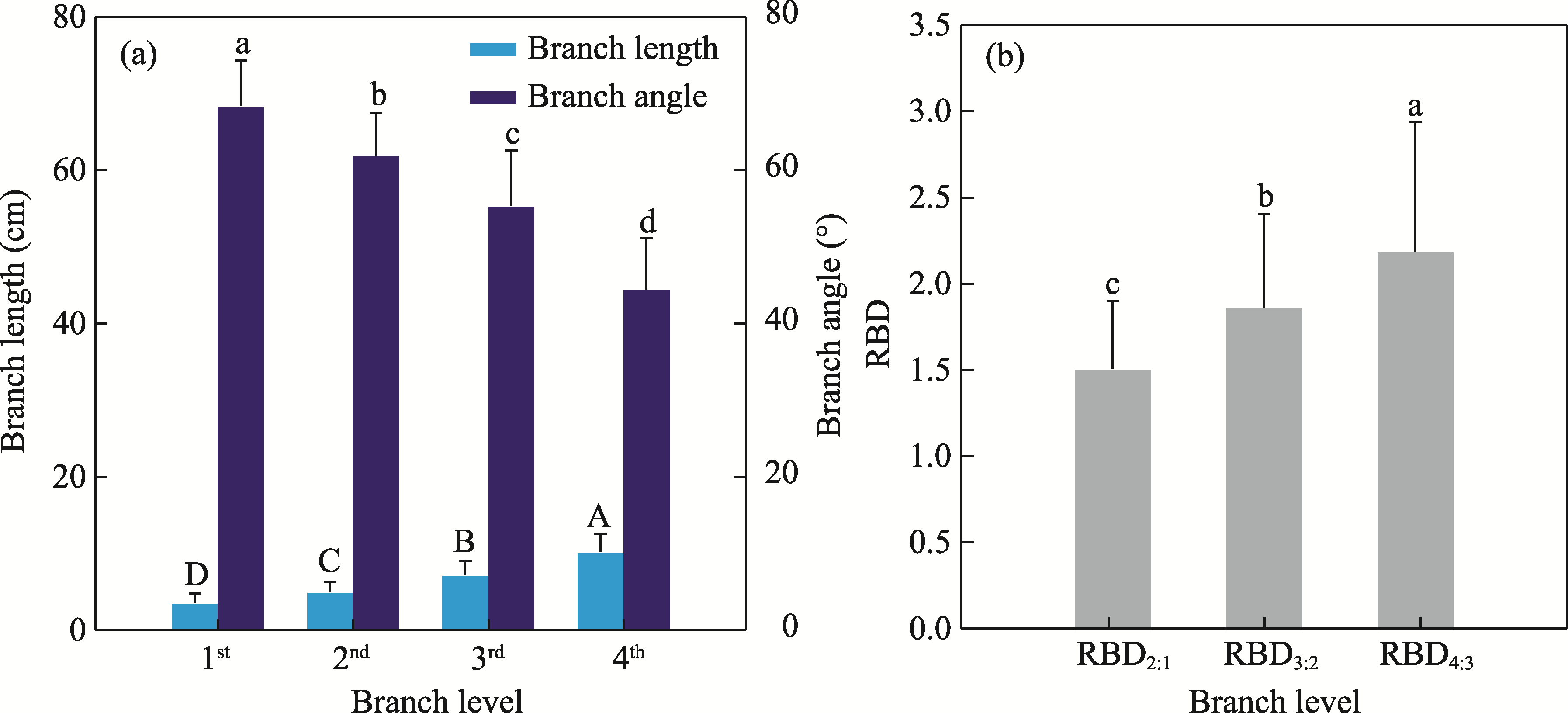
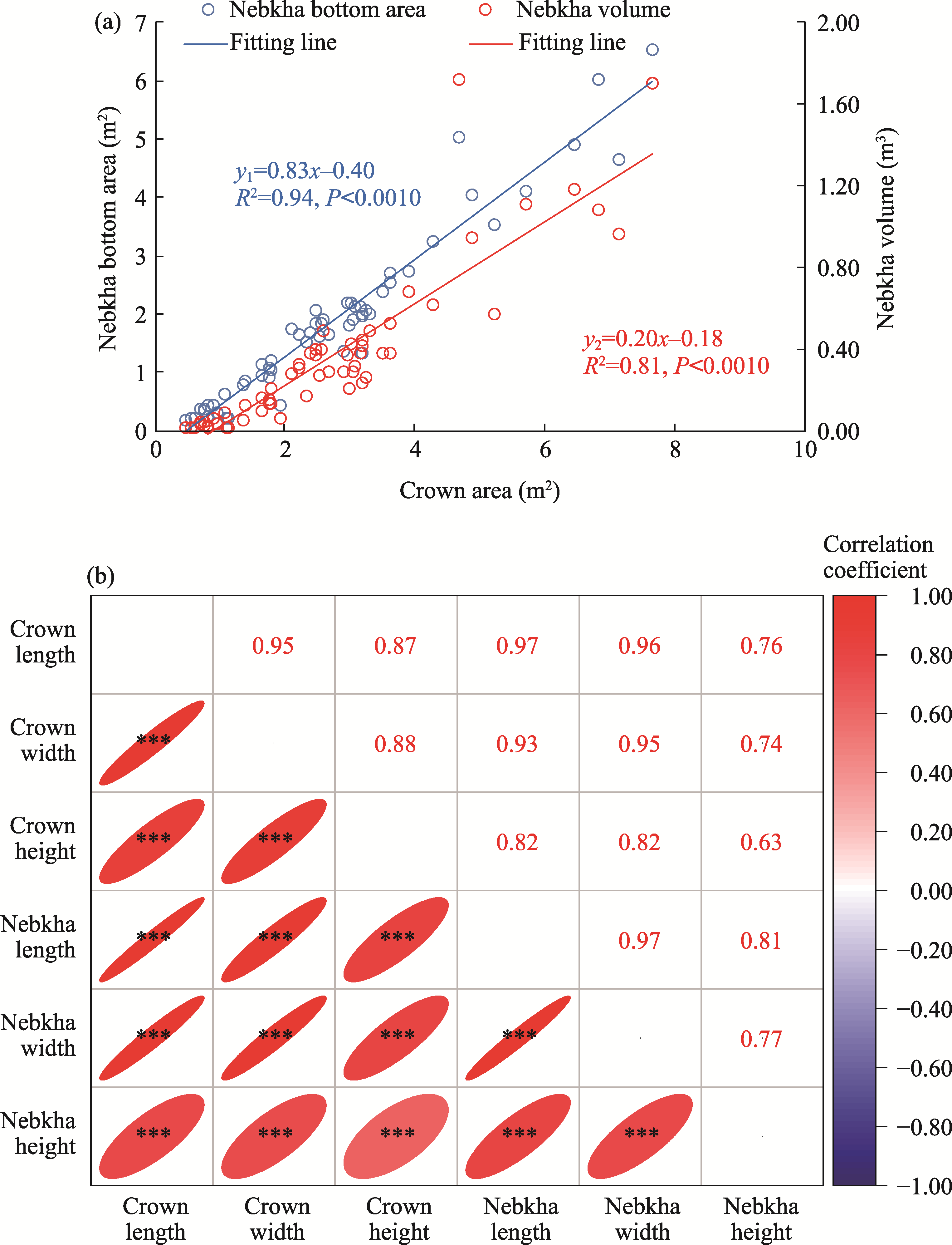
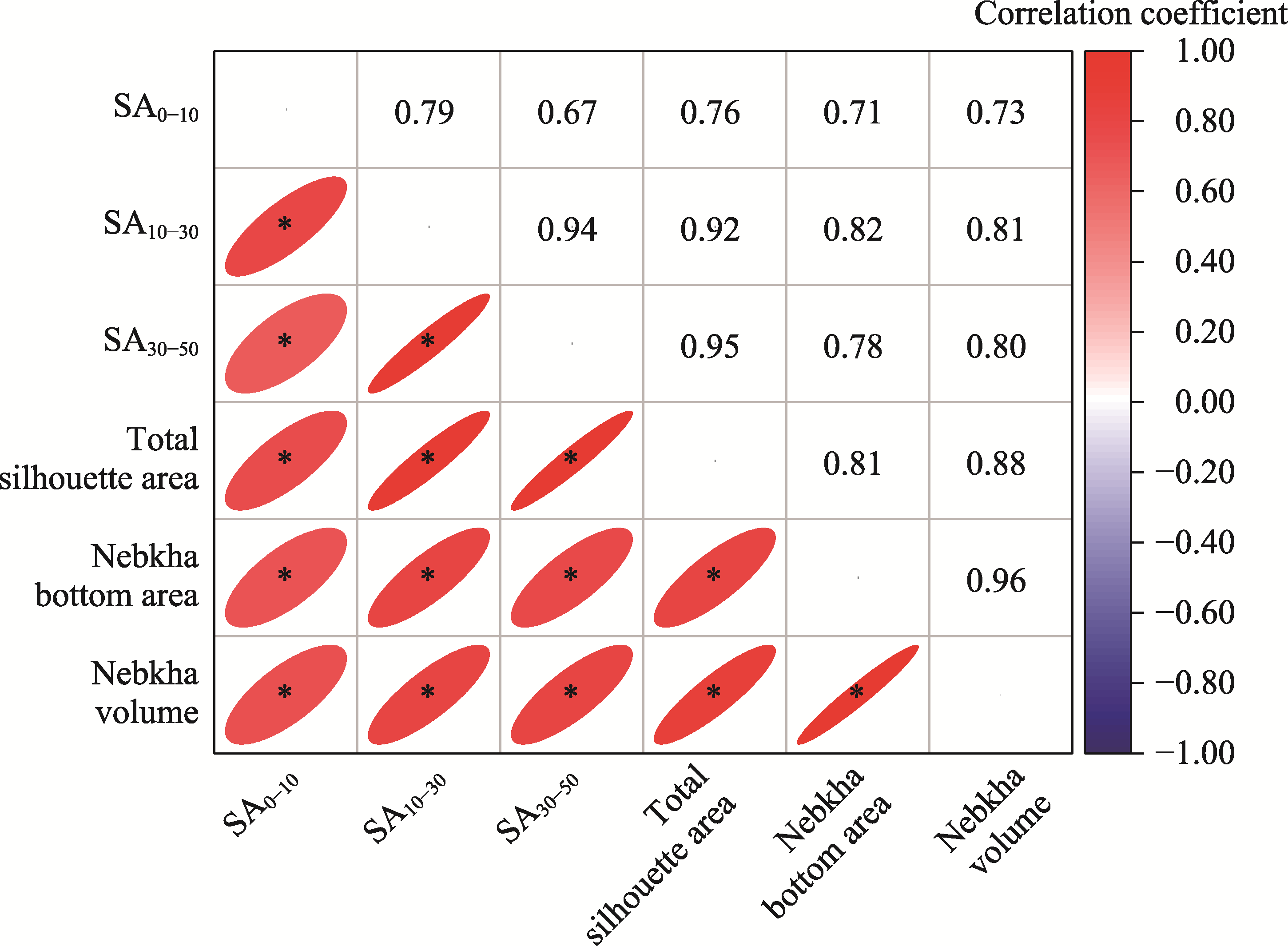
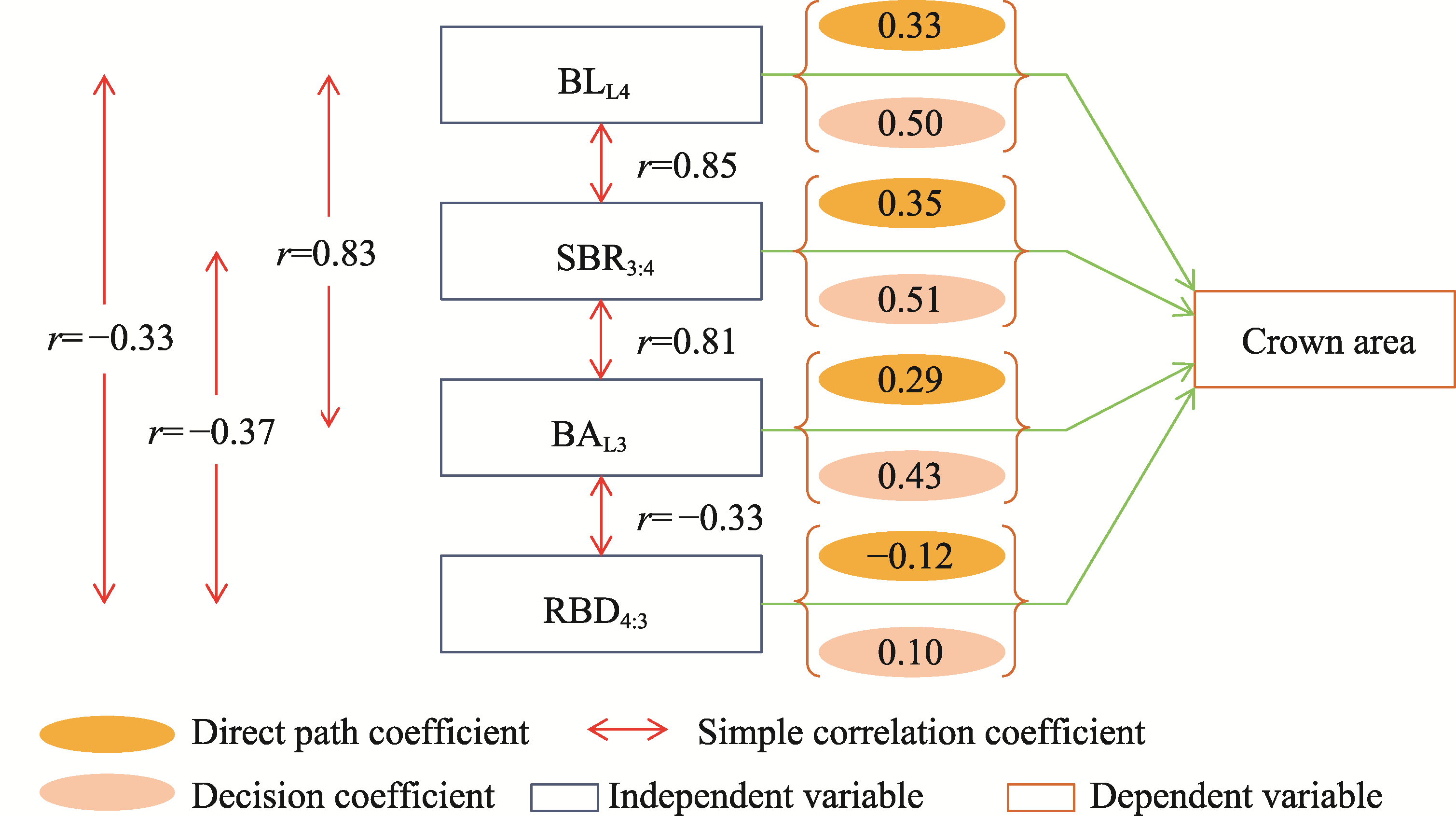
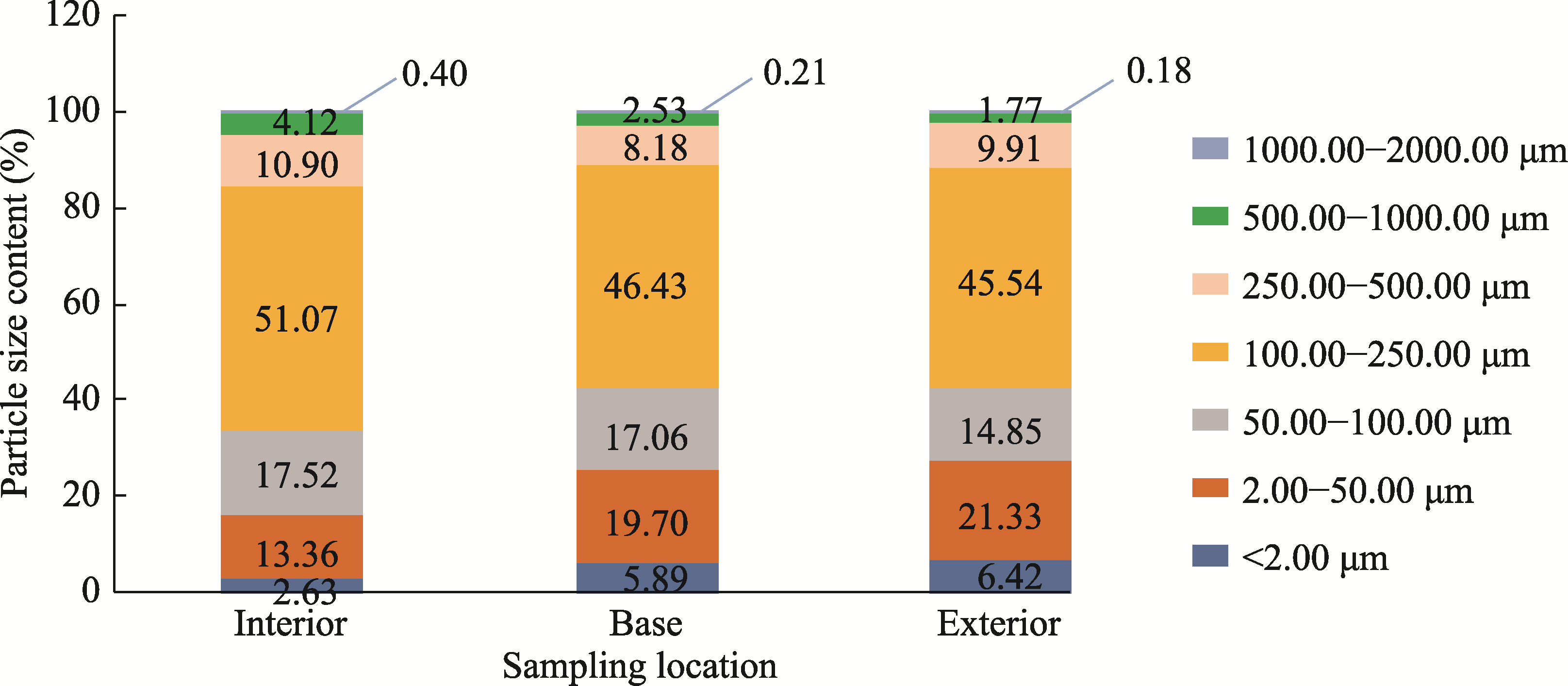
Abstract The formation of desert shrub sand piles (nebkhas) is attributed to the obstruction and subsequent deposition of migrating sand by the shrub itself. However, the relationship between sediment particle size distribution and shrub branch architecture remains inadequately understood. In August 2020, field investigations were conducted on Tetraena mongolica Maxim. shrubs in the Bayan Engger Desert Nature Reserve, located on the Ordos Plateau in Inner Mongolia Autonomous Region, China. Crown morphological parameters of T. mongolica shrubs and associated nebkhas were systematically measured alongside branch architectures. A one-way analysis of variance (ANOVA) was used to identify differences in branch architectures among various levels, while correlation analysis and model fitting were applied to establish the relationship between crown and nebkha morphological parameters. Path analysis was utilized to identify the key branch architectures that influence crown development. Furthermore, sediment redistribution characteristics of nebkhas were quantified, and principal component analysis combined with regression models was utilized to elucidate the contributions of key branch architectures and sensitive particle size fractions to nebkha deposition. Results indicated that the step-by-step branch ratio (SBR) initially increased from the lower branches to the outermost branches before subsequently decreasing. Additionally, branch angle significantly increased (P<0.0500), whereas both the branch length and the ratio of branch diameters (RBD) significantly decreased toward the exterior of the shrub (P<0.0500). Expansion of crown area significantly enhanced nebkha volume, demonstrating a strong linear relationship (P<0.0010). As the primary contact surface for trapping wind-blown sand, the silhouette area of the shrub initially increased and then decreased from bottom to top. Notably, the silhouette area of the 10-30 cm height layer played a crucial role in promoting nebkha volume expansion (P<0.0100). Path analysis further revealed that the key branch architectures promoting crown area expansion were the step-by-step branch ratio between the third-level and fourth-level branches (SBR3:4), followed by the fourth-level branch length (BLL4), the third-level branch angle (BAL3), and the ratio of branch diameters between the fourth-level and third-level branches (RBD4:3). Under the continuous interception of sediments by branches and leaves, the proportion of surface sediment with a particle size of 100.00-250.00 μm reached 51.07%, indicating a significant increase in fine-sized particles. Further analysis confirmed that SBR3:4, BLL4, BAL3, and sediments within the 50.00-100.00 μm particle size range were the primary contributors to nebkha deposition. These results demonstrate that the branch characteristics of T. mongolica shrubs near the ground surface promote fine sediment accumulation and nebkha development by regulating crown expansion. The findings reveal the unique adaptation mechanisms of rare and endangered plants in nebkha microhabitats and provide a scientific basis for ecological windbreak and sand-fixation projects in the desert transition zones of arid and semi-arid regions.
|
|
Received: 10 October 2025
Published: 31 May 2026
|
|
Corresponding Authors:
*DANG Xiaohong (E-mail: dangxiaohong1986@126.com)
|
| About author: Author contributions
Conceptualization: ZHAI Bo, DANG Xiaohong; Data curation: ZHAI Bo, DANG Xiaohong, LIU Xiangjie; Formal analysis: ZHAI Bo, LIU Jing, CHEN Xiaona; Funding acquisition: ZHAI Bo, DANG Xiaohong; Investigation: ZHAI Bo, DANG Xiaohong, LIU Jing, LIU Xiangjie, CHEN Xiaona, LIU Yajing; Methodology: ZHAI Bo, LIU Jing, LIU Xiangjie, CHEN Xiaona, LIU Yajing; Project administration: DANG Xiaohong; Resources: ZHAI Bo, DANG Xiaohong; Software: ZHAI Bo, LIU Jing, CHEN Xiaona, LIU Yajing; Supervision: DANG Xiaohong; Validation: ZHAI Bo, LIU Jing, LIU Xiangjie, CHEN Xiaona; Visualization: ZHAI Bo, LIU Xiangjie, LIU Yajing; Writing - original draft: ZHAI Bo; Writing - review and editing: ZHAI Bo, DANG Xiaohong, LIU Xiangjie, CHEN Xiaona. All authors approved the manuscript.
|
|
|
| [1] |
Araus J L, Kefauver S C, Díaz O V, et al. 2022. Crop phenotyping in a context of global change: What to measure and how to do it. Journal of Integrative Plant Biology, 64(2): 592-618.
doi: 10.1111/jipb.13191
|
|
|
| [2] |
Awogbemi C A, Alagbe S A, Oloda F S. 2022. On the path analysis techniques and decomposition of correlation coefficients. Asian Journal of Probability and Statistics, 20(4): 208-219.
|
|
|
| [3] |
Barthélémy D, Caraglio Y. 2007. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Annals of Botany, 99(3): 375-407.
doi: 10.1093/aob/mcl260
pmid: 17218346
|
|
|
| [4] |
Bazzaz F A. 1979. The physiological ecology of plant succession. Annual Review of Ecology, Evolution, and Systematics, 10(1): 351-371.
|
|
|
| [5] |
Borchert R, Slade N. 1981. Bifurcation ratios and the adaptive geometry of trees. Botanical Gazette, 142(3): 394-401.
doi: 10.1086/337238
|
|
|
| [6] |
Chambers S M. 2020. Novel methodologies to disentangle plant-environment interactions. Applications in Plant Sciences, 8(2): e11324, doi: 10.1002/aps3.11324.
|
|
|
| [7] |
Chang Z F, Zhang J H, Shi X G, et al. 2017. Initial research on the relationship between sand-mound formation and the layered silhouette of desert plants. Acta Ecologica Sinica, 37(21): 7351-7358. (in Chinese)
|
|
|
| [8] |
Cheng H, Zhang K D, Liu C C, et al. 2018. Wind tunnel study of airflow recovery on the lee side of single plants. Agricultural and Forest Meteorology, 263: 362-372.
doi: 10.1016/j.agrformet.2018.08.025
|
|
|
| [9] |
Ciancio M E, Guida-Johnson B, Zuleta G A, et al. 2022. Rehabilitation planning in an oilfield in the Monte Austral: Mapping sand‐sized sediment availability and assessing its effect on microtopography rehabilitation. Earth Surface Processes and Landforms, 47(13): 3133-3146.
doi: 10.1002/esp.v47.13
|
|
|
| [10] |
Clark C B, Ma J X. 2023. The genetic basis of shoot architecture in soybean. Molecular Breeding, 43: 55, doi: 10.1007/s11032-023-01391-3.
|
|
|
| [11] |
Costas S, Sousa L B D, Gallego-Fernández J B, et al. 2024. Foredune initiation and early development through biophysical interactions. Science of The Total Environment, 940: 173548, doi: 10.1016/j.scitotenv.2024.173548.
|
|
|
| [12] |
Dos Santos P, Brilhante M A, Messerschmid T F E, et al. 2022. Plant growth forms dictate adaptations to the local climate. Frontiers in Plant Science, 13: 1023595, doi: 10.3389/fpls.2022.1023595.
|
|
|
| [13] |
El-Sheikh M A, Abbadi G A, Bianco P M, et al. 2010. Vegetation ecology of phytogenic hillocks (nabkhas) in coastal habitats of Jal Az-Zor National Park, Kuwait: Role of patches and edaphic factors. Flora, 205(12): 832-840.
doi: 10.1016/j.flora.2010.01.002
|
|
|
| [14] |
Enquist B J, Brown J H, West G B. 1998. Allometric scaling of plant energetics and population density. Nature, 395: 163-165.
doi: 10.1038/25977
|
|
|
| [15] |
Fisher J B, Honda H. 1979. Branch geometry and effective leaf area: a study of Terminalia-branching pattern. 1. Theoretical trees. American Journal of Botany, 66(6): 633-644.
|
|
|
| [16] |
Goudie S. 2022. Nebkhas: An essay in aeolian biogeomorphology. Aeolian Research, 54: 100772, doi: 10.1016/j.aeolia.2022.100772.
|
|
|
| [17] |
Guan Q Y, Zhang J D, Wang L J, et al. 2013. Discussion of the relationship between dustfall grain size and the desert border, taking the southern border of the Tengger Desert and the southern dust deposit area as an example. Palaeogeography, Palaeoclimatology, Palaeoecology, 386(17): 1-7.
doi: 10.1016/j.palaeo.2013.01.017
|
|
|
| [18] |
Haynes M, Fanni R. 2025. Development of a laboratory wind tunnel to test tailings erosion mechanisms. In: Knutsson S, Fourie A B, Tibbett M. Mine Closure 2025: Proceedings of the 18th International Conference on Mine Closure, Australian Centre for Geomechanics. Perth, Australia. doi: 10.36487/ACG_repo/2515_90.
|
|
|
| [19] |
He M Z, Wang H, Zhang J G, et al. 2005. Classification of the branching architectures of the desert plants in Minqin County. Acta Botanica Boreali-Occidentalia Sinica, 25(9): 1827-1832. (in Chinese)
|
|
|
| [20] |
Hesp P A. 1981. The formation of shadow dunes. Journal of Sedimentary Research, 51(1): 101-112.
|
|
|
| [21] |
Hildebrand M, Perles-Garcia M D, Kunz M, et al. 2021. Tree-tree interactions and crown complementarity: the role of functional diversity and branch traits for canopy packing. Basic and Applied Ecology, 50: 217-227.
doi: 10.1016/j.baae.2020.12.003
|
|
|
| [22] |
Honda H. 1971. Description of the form of trees by the parameters of the tree-like body: Effects of the branching angle and the branch length on the shape of the tree-like body. Journal of Theoretical Biology, 31(2): 331-334.
doi: 10.1016/0022-5193(71)90191-3
pmid: 5557081
|
|
|
| [23] |
Hu G Y, Dong Z B, Lu J F, et al. 2021. Spatial pattern of aeolian desertification and its causes in the Yellow River catchment. Journal of Desert Research, 41(4): 213-224. (in Chinese)
doi: 10.7522/j.issn.1000-694X.2021.00084
|
|
|
| [24] |
Jin Z, Dong Y S, Qi Y C, et al. 2013. Characterizing variations in soil particle-size distribution along a grass-desert shrub transition in the Ordos Plateau of Inner Mongolia, China. Land Degradation & Development, 24(2): 141-146.
doi: 10.1002/ldr.v24.2
|
|
|
| [25] |
Kidron G J, Zohar M. 2016. Factors controlling the formation of coppice dunes (nebkhas) in the Negev Desert. Earth Surface Processes and Landforms, 41(7): 918-927.
doi: 10.1002/esp.v41.7
|
|
|
| [26] |
Koller A, Kunz M, Perles-Garcia M D et al. 2025. 3D structural complexity of forest stands is determined by the magnitude of inner and outer crown structural attributes of individual trees. Agricultural and Forest Meteorology, 363: 110424, doi: 10.1016/j.agrformet.2025.110424.
|
|
|
| [27] |
Lancaster N, Baas A. 2015. Influence of vegetation cover on sand transport by wind: Field studies at Owens Lake, California. Earth Surface Processes and Landforms, 23(1): 69-82.
doi: 10.1002/(ISSN)1096-9837
|
|
|
| [28] |
Leopold L B. 1971. Trees and streams: the efficiency of branching patterns. Journal of Theoretical Biology, 31(2): 339-354.
pmid: 5557082
|
|
|
| [29] |
Li J C, Yao Q, Wang Y, et al. 2019. Grain-size characteristics of surface sediments of nebkhas at the southern margin of the Mu Us dune field, China. CATENA, 183: 104210, doi: 10.1016/j.catena.2019.104210.
|
|
|
| [30] |
Li J R, Ravi S. 2018. Interactions among hydrological-aeolian processes and vegetation determine grain-size distribution of sediments in a semi-arid coppice dune (nebkha) system. Journal of Arid Environments, 154(7): 24-33.
doi: 10.1016/j.jaridenv.2018.03.011
|
|
|
| [31] |
Li S H, Mason J A, Xu Y H, et al. 2021. Biogeomorphology of nebkhas in the Mu Us dune field, north-central China: Chronological and morphological results. Geomorphology, 394: 107979, doi: 10.1016/j.geomorph.2021.107979.
|
|
|
| [32] |
Liu J W, Li Z Z, Wu S L, et al. 2009. The spatial heterogeneity of morphologic feature of Nitraria nebkhas around Ebinur Lake‚ Xinjiang. Journal of Desert Research, 29(4): 628-635. (in Chinese)
|
|
|
| [33] |
Liu Z K, Wang C W, Yang X J, et al. 2023. The relationship and influencing factors between endangered plant Tetraena mongolica and soil microorganisms in West Ordos desert ecosystem, northern China. Plants, 12(5): 1048, doi: 10.3390/plants12051048.
|
|
|
| [34] |
Lu Y, Zhang B R, Zhang M, et al. 2023. Relict plants are better able to adapt to climate change: Evidence from desert shrub communities. Plants, 12(23): 4065, doi: 10.3390/plants12234065.
|
|
|
| [35] |
Niklas K J, Cobb E D. 2010. Ontogenetic changes in the numbers of short- vs. long-shoots account for decreasing specific leaf area in Acer rubrum (Aceraceae) as trees increase in size. American Journal of Botany, 97(1): 27-37.
doi: 10.3732/ajb.0900249
pmid: 21622364
|
|
|
| [36] |
Osada N. 2006. Crown development in a pioneer tree, rhus trichocarpa, in relation to the structure and growth of individual branches. New Phytologist, 172(4): 667-678.
pmid: 17096793
|
|
|
| [37] |
Pang Y J, Wu B, Jia X H, et al. 2022. Wind-proof and sand-fixing effects of Artemisia ordosica with different coverages in the Mu Us Sandy Land, northern China. Journal of Arid Land, 14(8): 877-893.
doi: 10.1007/s40333-022-0070-4
|
|
|
| [38] |
Qu Z Q, Liu L Y, Lü Y L. 2011. Psammophyte architecture and its relations with anti-wind erosion capability: a review. Chinese Journal of Ecology, 30(2): 357-362. (in Chinese)
|
|
|
| [39] |
Qu Z Q, Li Z G, Hu L X, et al. 2022. The branching architecture of Artemisia ordosica and its resistance to wind erosion. Frontiers in Environmental Science, 10: 960969, doi: 10.3389/fenvs.2022.960969.
|
|
|
| [40] |
Savinov I A. 2020. Architectural analysis of representatives of the Celastrales order: structure and rhythm of shoot development in connection with adaptations of species to different environmental conditions. Contemporary Problems of Ecology, 13: 300-308.
doi: 10.1134/S1995425520030099
|
|
|
| [41] |
Steingraeber D A, Kascht L J, Franck D H. 1979. Variation of shoot morphology and bifurcation ratio in sugar maple (Acer saccharum) saplings. American Journal of Botany, 66(4): 441-445.
doi: 10.1002/ajb2.1979.66.issue-4
|
|
|
| [42] |
Steingraeber D A, Waller D M. 1986. Non-stationarity of tree branching patterns and bifurcation ratios. Proceedings of the Royal Society B: Biological Sciences, 228(1251): 187-194.
|
|
|
| [43] |
Sweeney M R, Lu H Y, Cui M C, et al. 2016. Sand dunes as potential sources of dust in northern China. Science China Earth Sciences, 59: 760-769.
doi: 10.1007/s11430-015-5246-8
|
|
|
| [44] |
Tengberg A. 1995. Nebkha dunes as indicators of wind erosion and land degradation in the Sahel zone of Burkina Faso. Journal of Arid Environments, 30(3): 265-282.
doi: 10.1016/S0140-1963(05)80002-3
|
|
|
| [45] |
Valladares F, Gianoli E, Gómez J M. 2007. Ecological limits to plant phenotypic plasticity. New Phytologist, 176(4): 749-763.
doi: 10.1111/j.1469-8137.2007.02275.x
pmid: 17997761
|
|
|
| [46] |
White J. 1979. The plant as a metapopulation. Annual Review of Ecology, Evolution, and Systematics, 10(1): 109-145.
|
|
|
| [47] |
Whitney G G. 1976. The bifurcation ratio as an indicator of adaptive strategy in woody plant species. Bulletin of the Torrey Botanical Club, 103(2): 67-72.
doi: 10.2307/2484833
|
|
|
| [48] |
Wolfe S A, Nickling W G. 1993. The protective role of sparse vegetation in wind erosion. Progress in Physical Geography, 17(1): 50-68.
|
|
|
| [49] |
Wu W Y, Zhang D S, Tian L H, et al. 2023. Morphological change and migration of revegetated dunes in the Ketu Sandy Land of the Qinghai Lake, China. Journal of Arid Land, 15(7): 827-841.
doi: 10.1007/s40333-023-0021-8
|
|
|
| [50] |
Xu D L, Yu X W, Chen J, et al. 2023. Arbuscular mycorrhizae fungi diversity in the root-rhizosphere-soil of Tetraena mongolica, Sarcozygium xanthoxylon, and Nitraria tangutorum Bobr in western Ordos, China. Agronomy, 13(6): 1485, doi: 10.3390/agronomy13061485.
|
|
|
| [51] |
Xu X J, Chen N M, Feng J C, et al. 2020. Comparative analyses of leaf cuticular lipids of two succulent xerophytes of the Ordos Plateau (Gobi Desert), Tetraena mongolica Maxim and Zygophyllum xanthoxylum (Bunge) Engl. Environmental and Experimental Botany, 177: 104129, doi: 10.1016/j.envexpbot.2020.104129.
|
|
|
| [52] |
Yagi T. 2006. Relationships between shoot size and branching patterns in 10 broad-leaved tall tree species in a Japanese cool-temperate forest. Canadian Journal of Botany, 84(12): 1894-1907.
doi: 10.1139/b06-138
|
|
|
| [53] |
Yan P, Deng Y H, An S J, et al. 2025. Training systems affect spatial distribution of Korla fragrant pear (Pyrus sinkiangensis Yu) fruits by altering canopy structure and light distribution. Frontiers in Plant Science, 16: 1615019, doi: 10.3389/fpls.2025.1615019.
|
|
|
| [54] |
Yang L W, Zhu S, Xu J. 2022. Roles of auxin in the inhibition of shoot branching in 'Dugan' fir. Tree Physiology, 42(7): 1411-1431.
doi: 10.1093/treephys/tpac008
|
|
|
| [55] |
Zhai B. 2022. The influence of the shrub configuration features of Tetraena mongolica Maxim on the aeolian processes and sedimentary characteristics. PhD Dissertation. Hohhot: Inner Mongolia Agricultural University. (in Chinese)
|
|
|
| [56] |
Zhang Z C, Han L Y, Pan K J, et al. 2020. Nebkha dune morphology in the gobi deserts of northern China and potential implications for dust emission. Sedimentology, 67(7): 3769-3782.
doi: 10.1111/sed.v67.7
|
|
|
| [57] |
Zhao X H, Miao Z, Li F R, et al. 2025. Unraveling the individual and interactive effects of climate and competition on branch growth dynamics in Pinus koraiensis in Northeast China. Frontiers in Plant Science, 16: 1545892, doi: 10.3389/fpls.2025.1545892.
|
|
|
| [58] |
Zhu G N, Nong H J, Fang S Y, et al. 2024. Arbuscular mycorrhizal symbiosis reshapes the drought adaptation strategies of a dominant sand-fixation shrub species in northern China. Science of The Total Environment, 955: 177135, doi: 10.1016/j.scitotenv.2024.177135.
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|