| Research article |
|

|
|
|
| Root system architecture and its scaling relationships of Reaumuria soongorica in Alxa steppe desert, Northwest China |
MA Xiongzhong1,2, WANG Xinping3,*( ), XIONG Weihong1,2 ), XIONG Weihong1,2 |
1School of Geographical Science and Planning, Nanning Normal University, Nanning 530001, China
2Key Laboratory of Environment Change and Resources Use in Beibu Gulf, Ministry of Education, Nanning Normal University, Nanning 530001, China
3Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China |
|
|
|
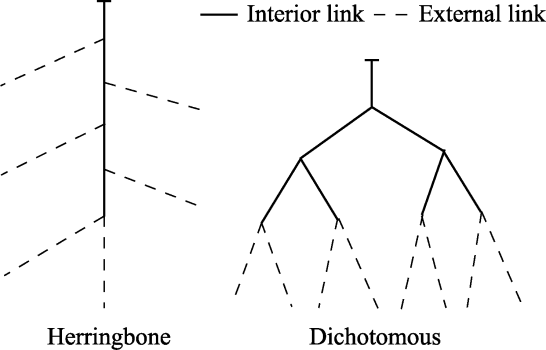
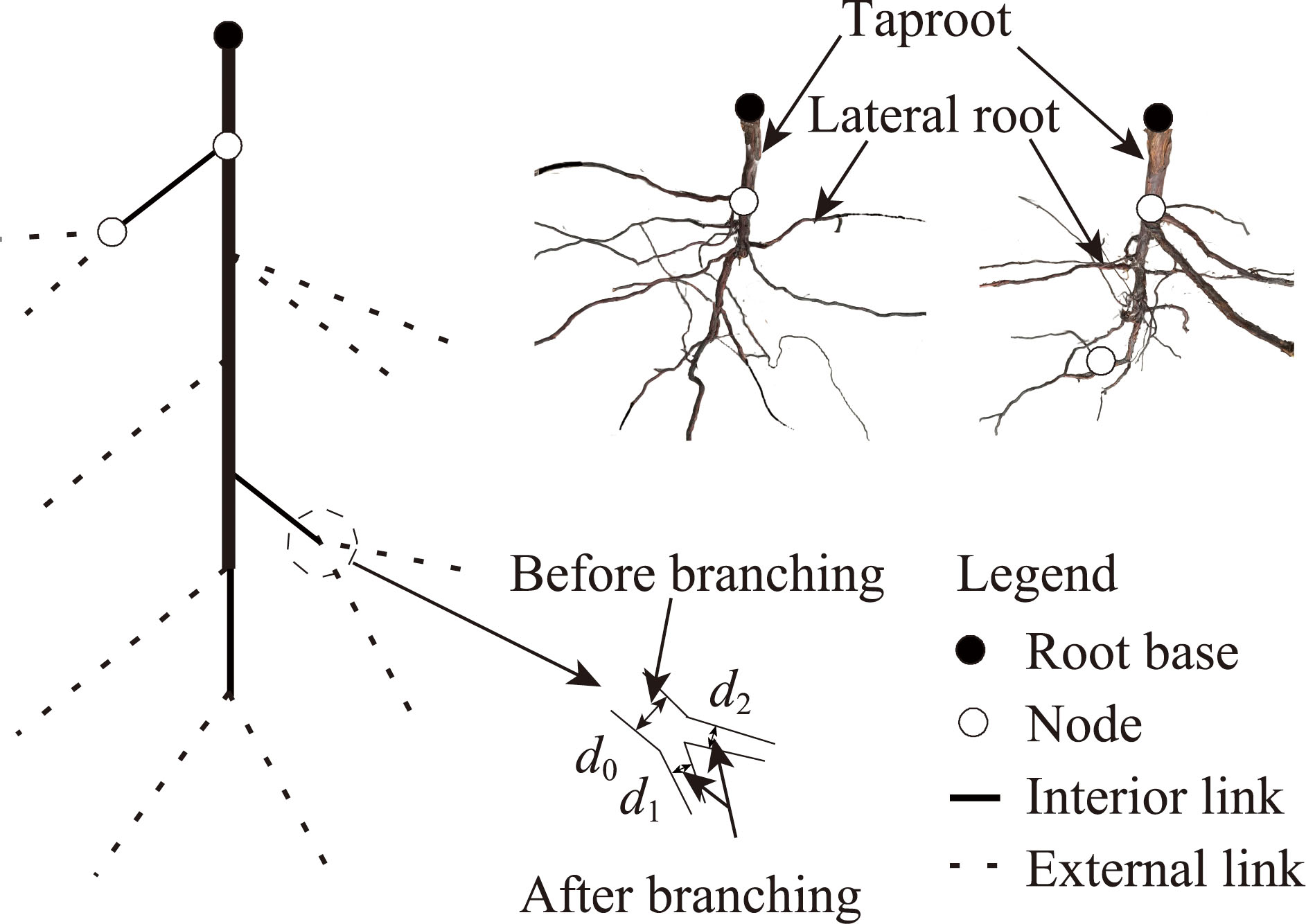
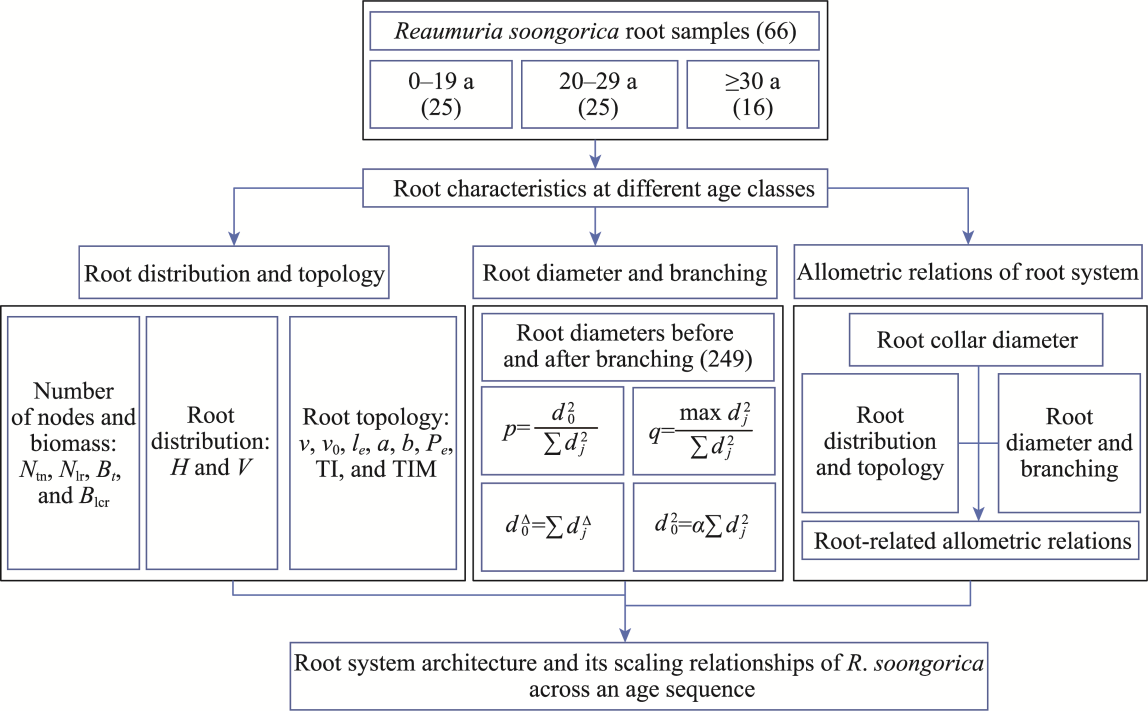
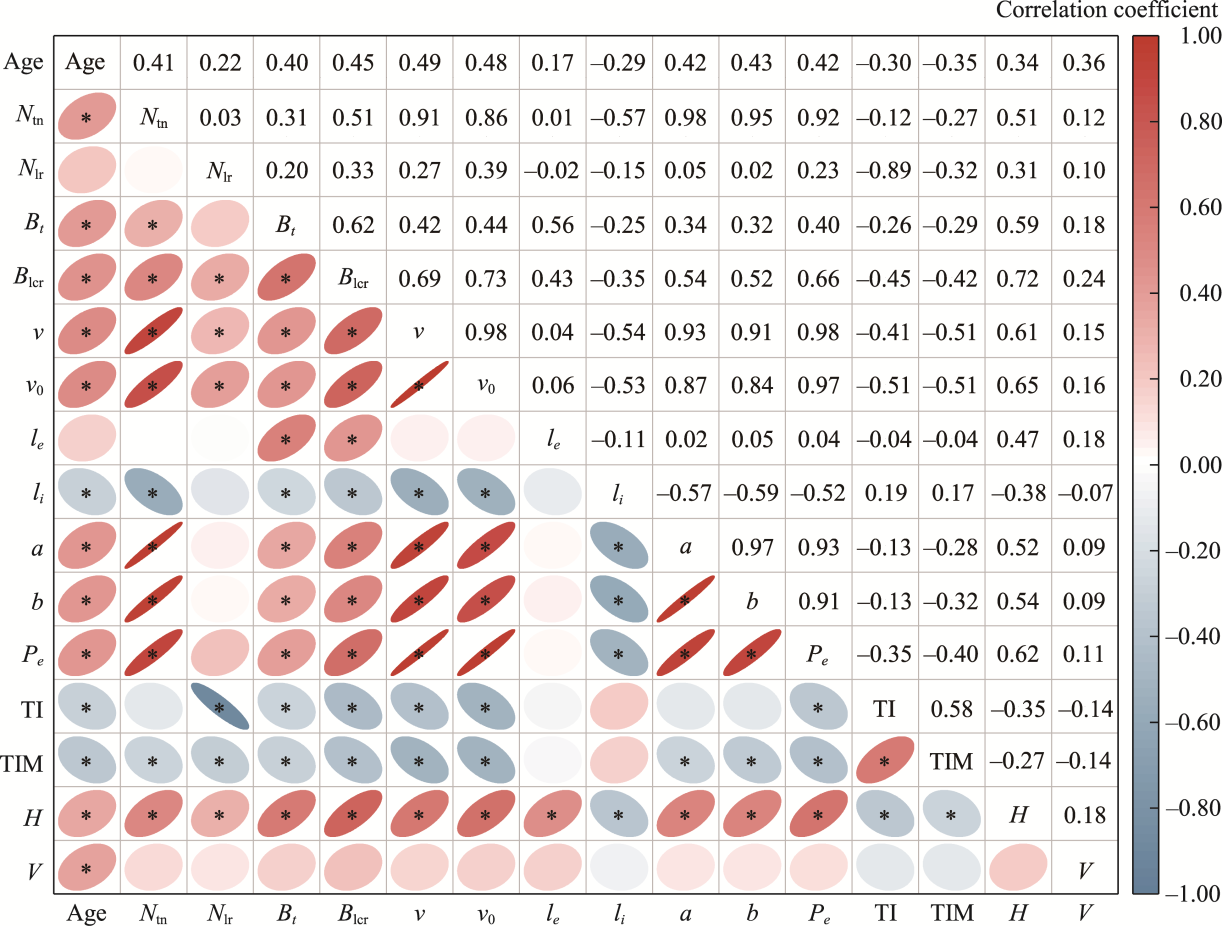
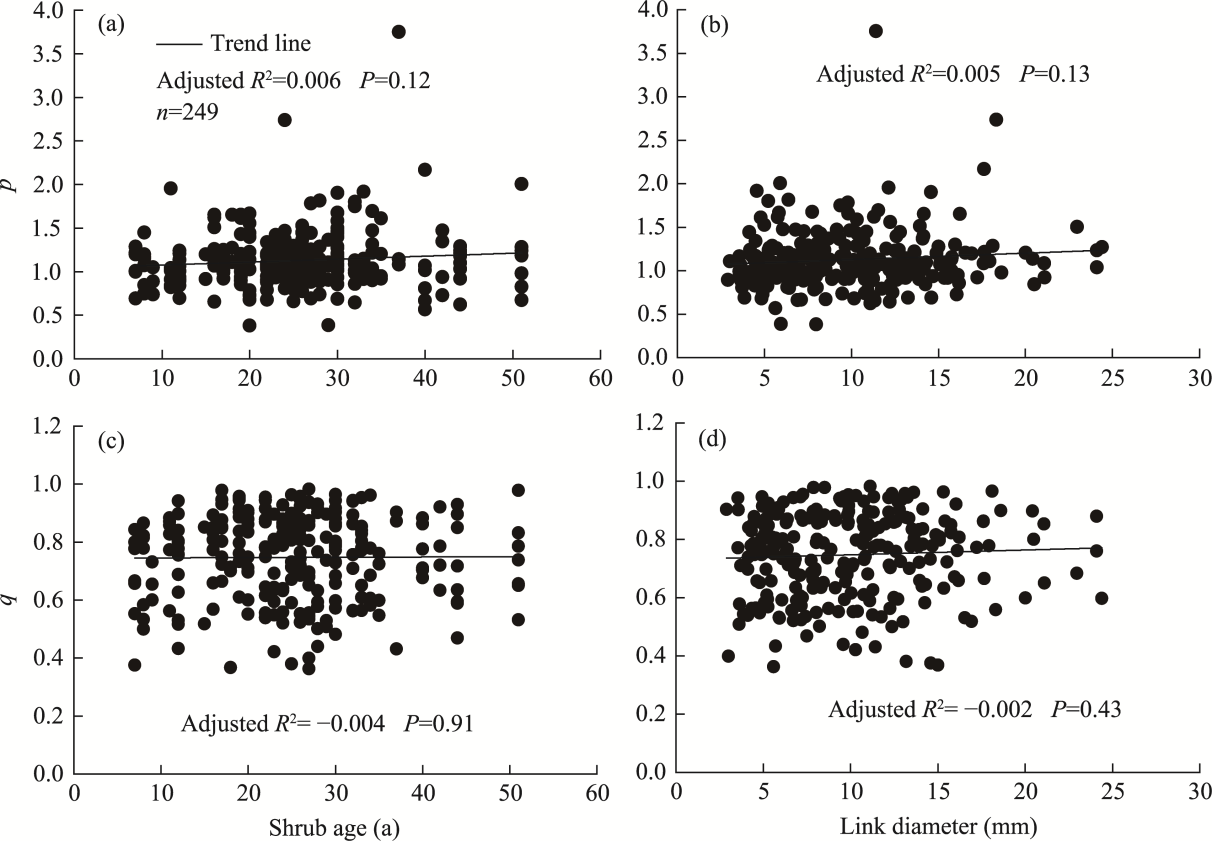
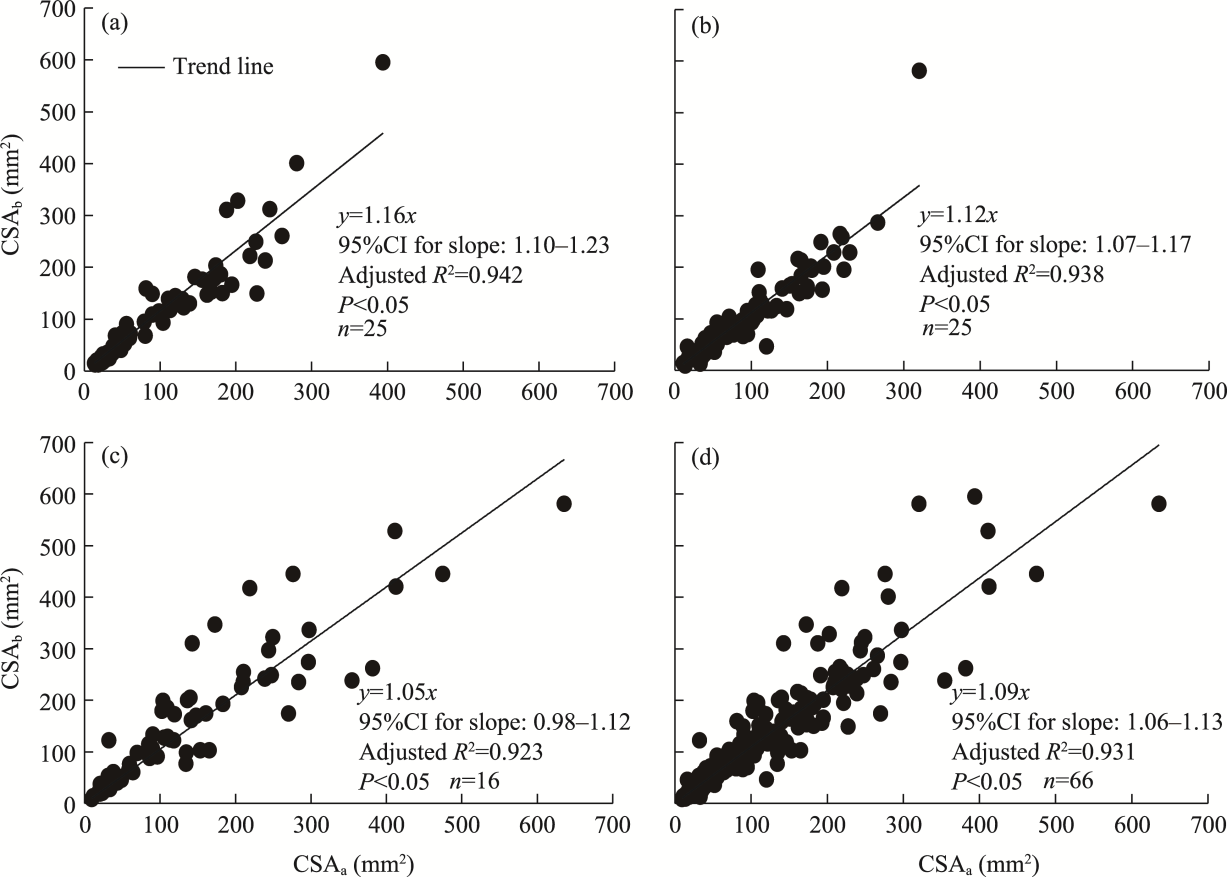
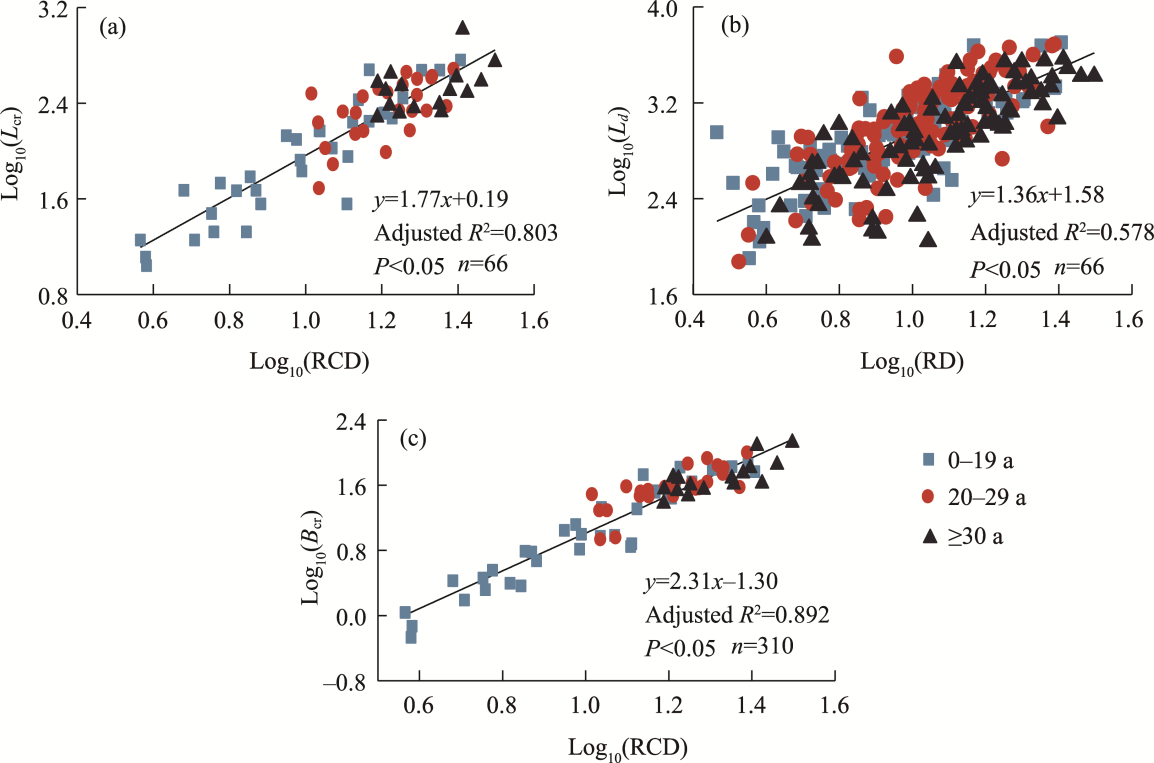
Abstract Root system architecture has often been overlooked in plant research despite its critical role in plant adaptation to environmental conditions. This study focused on the root system architecture of the desert shrub Reaumuria soongorica in the Alxa steppe desert, Northwest China. Plant samples were collected during May-September 2019. Using excavation methods, in situ measurements, and root scanning techniques, we analyzed the root distribution, topology, and branching patterns of R. soongorica across an age sequence of 7-51 a. Additionally, we investigated the allometric relationships of root collar diameter with total coarse root length, biomass, and topological parameters. The results showed that the roots of R. soongorica were predominantly concentrated in shallow soil layers (10-50 cm), with lateral root branching and biomass allocation increasing with shrub age. The root topology exhibited a herringbone-like structure, with average topological and modified topological indices of 0.89 and 0.96, respectively, both of which adjusted with shrub age. The root system displayed a self-similar branching pattern, maintaining a constant cross-sectional area ratio of 1.13 before and after branching, deviating from the area-preserving rule. These adaptive traits allow R. soongorica to efficiently expand its nutrient acquisition zone, minimize internal competition, and optimize resource uptake from the upper soil layers. Furthermore, significant linear relationships were observed between log10-transformed root collar diameter and log10-transformed total coarse root length, biomass, and topological parameters. These findings advance non-destructive approaches for studying root characteristics and contribute to the development of root-related models. Besides, this study provides new insights into the adaptive strategies of R. soongorica under extreme drought conditions, offering valuable guidance for species selection and cultivation in desert restoration efforts.
|
|
Received: 23 July 2024
Published: 28 February 2025
|
|
Corresponding Authors:
*WANG Xinping (E-mail: xpwang@lzb.ac.cn)
|
|
|
| [1] |
Anfodillo T, Carraro V, Carrer M, et al. 2006. Convergent tapering of xylem conduits in different woody species. New Phytologist, 169(2): 279-290.
pmid: 16411931
|
|
|
| [2] |
Bai X, Zhao C Z, Kang M P. 2021. Relationship between root forks and branch angle of Tamarix ramosissima at different stand ages in oasis of Jinta County. Acta Ecologica Sinica, 41(5): 1878-1884. (in Chinese)
|
|
|
| [3] |
Bucci S J, Scholz F G, Peschiutta M L, et al. 2013. The stem xylem of Patagonian shrubs operates far from the point of catastrophic dysfunction and is additionally protected from drought-induced embolism by leaves and roots. Plant, Cell & Environment, 36(12): 2163-2174.
|
|
|
| [4] |
Chen X Y, Chen Y J, Zhang W, et al. 2023. Response characteristics of root to moisture change at seedling stage of Kengyilia hirsuta. Frontiers in Plant Science, 13: 1052791, doi: 10.3389/fpls.2022.1052791.
|
|
|
| [5] |
Dhief A, Abdellaoui R, Tarhouni M, et al. 2011. Root and aboveground growth of rhizotron-grown seedlings of three Tunisian desert Calligonum species under water deficit. Canadian Journal of Soil Science, 91(1): 15-27.
|
|
|
| [6] |
Fitter A H. 1987. An architectural approach to the comparative ecology of plant root system. New Phytologist, 106(S1): 61-77.
|
|
|
| [7] |
He G Z, Chen Y N, Chen Y P, et al. 2016. Adaptive strategy of Tamarix spp root architecture in arid environment. Journal of Beijing Normal University (Natural Science), 52(3): 277-282. (in Chinese)
|
|
|
| [8] |
Karlova R, Boer D, Hayes S, et al. 2021. Root plasticity under abiotic stress. Plant Physiology, 187(3): 1057-1070.
doi: 10.1093/plphys/kiab392
pmid: 34734279
|
|
|
| [9] |
Kirschner G K, Xiao T T, Blilou I. 2021. Rooting in the desert: A developmental overview on desert plants. Genes, 12(5): 709, doi: 10.3390/genes12050709.
|
|
|
| [10] |
Li C J, Guo J H, Zeng F J, et al. 2015. Shoot and root architectural variance and adaptability of Tamarix ramosissima in different ages. Journal of Desert Research, 35(2): 365-372. (in Chinese)
|
|
|
| [11] |
Li C J, Han H, Ablimiti M, et al. 2022a. Morphological and physiological responses of desert plants to drought stress in a man-made landscape of the Taklimakan desert shelter belt. Ecological Indicators, 140: 109037, doi: 10.1016/j.ecolind.2022.109037.
|
|
|
| [12] |
Li X L, Dang X H, Gao Y, et al. 2022b. Response mechanisms of adventitious root architectural characteristics of Nitraria tangutorum shrubs to soil nutrients in Nabkha. Plants, 11(23): 3218, doi: 10.3390/plants11233218.
|
|
|
| [13] |
Lynch J P. 2013. Steep, cheap and deep: an ideotype to optimize water and N acquisition by maize root systems. Annals of Botany, 112(2): 347-357.
doi: 10.1093/aob/mcs293
pmid: 23328767
|
|
|
| [14] |
Lynch J P. 2018. Rightsizing root phenotypes for drought resistance. Journal of Experimental Botany, 69(13): 3279-3292.
doi: 10.1093/jxb/ery048
pmid: 29471525
|
|
|
| [15] |
Ma X Z, Wang X P. 2020. Biomass partitioning and allometric relations of the Reaumuria soongorica shrub in Alxa steppe desert in NW China. Forest Ecology and Management, 468: 118178, doi: 10.1016/j.foreco.2020.118178.
|
|
|
| [16] |
Ma X Z, Wang X P, Jin Y X, et al. 2021. Fine root production, turnover of Reaumuria songarica and soil carbon and nitrogen in Alxa steppe desert of NW China. Restoration Ecology, 29(6): e13362, doi: 10.1111/rec.13362.
|
|
|
| [17] |
Ma X Z, Wang X P, Xiong W H. 2024. Fine-root turnover and leaf litterfall of Salsola passerina facilitate soil restoration in Alxa steppe desert of northwest China. Restoration Ecology, 32(6): e14040, doi: 10.1111/rec.14040.
|
|
|
| [18] |
Magalhães T M, Seifert T. 2015. Below- and aboveground architecture of Androstachys johnsonii prain: topological analysis of the root and shoot systems. Plant and Soil, 394(1): 257-269.
|
|
|
| [19] |
Nikolova P S, Blaschke H, Matyssek R, et al. 2009. Combined application of computer tomography and light microscopy for analysis of conductive xylem area in coarse roots of European beech and Norway spruce. European Journal of Forest Research, 128: 145-153.
|
|
|
| [20] |
Ober E S, Alahmad S, Cockram J, et al. 2021. Wheat root systems as a breeding target for climate resilience. Theoretical and Applied Genetics, 134(6): 1645-1662.
doi: 10.1007/s00122-021-03819-w
pmid: 33900415
|
|
|
| [21] |
Omary M, Gil-Yarom N, Chen Y, et al. 2022. A conserved superlocus regulates above- and belowground root initiation. Science, 375(6584): eabf4368, doi: 10.1126/science.abf4368.
|
|
|
| [22] |
Oppelt A L, Kurth W, Dzierzon H, et al. 2000. Structure and fractal dimensions of root systems of four co-occurring fruit tree species from Botswana. Annals of Forest Science, 57(5-6): 463-475.
|
|
|
| [23] |
Pan Y X, Wang X P, Zhang Y F, et al. 2018. Dew formation characteristics at annual and daily scale in xerophyte shrub plantations at Southeast margin of Tengger desert, Northern China. Ecohydrology, 11(5): e1968, doi: 10.1002/eco.1968.
|
|
|
| [24] |
Preti F, Dani A, Noto L V, et al. 2022. On the Leonardo's rule for the assessment of root profile. Ecological Engineering, 179: 106620, doi: 10.1016/j.ecoleng.2022.106620.
|
|
|
| [25] |
Price C A, Drake P, Veneklaas E J, et al. 2022. Flow similarity, stochastic branching, and quarter-power scaling in plants. Plant Physiology, 190(3): 1854-1865.
|
|
|
| [26] |
Ranjan A, Sinha R, Singla-Pareek S L, et al. 2022. Shaping the root system architecture in plants for adaptation to drought stress. Physiologia Plantarum, 174(2): e13651, doi: 10.1111/ppl.13651.
|
|
|
| [27] |
Richter J P. 1970. The Notebooks of Leonardo Da Vinci. New York: Dover.
|
|
|
| [28] |
Salas E, Ozier-Lafontaine H, Nygren P. 2004. A fractal root model applied for estimating the root biomass and architecture in two tropical legume tree species. Annals of Forest Science, 61(4): 337-345.
|
|
|
| [29] |
Shan L S, Li Y, Ren W, et al. 2013. Root architecture of two desert plants in central Hexi Corridor of Northwest China. Chinese Journal of Applied Ecology, 24(1): 25-31. (in Chinese)
pmid: 23717986
|
|
|
| [30] |
Shan L S, Su M, Zhang Z Z, et al. 2018. Vertical distribution pattern of mixed root systems of desert plants Reaumuria soongarica and Salsola passerina under different environmental gradients. Chinese Journal of Plant Ecology, 42(4): 475-486. (in Chinese)
|
|
|
| [31] |
Shelden M C, Munns R. 2023. Crop root system plasticity for improved yields in saline soils. Frontiers in Plant Science, 14: 1120583, doi: 10.3389/fpls.2023.1120583.
|
|
|
| [32] |
Sorgonà A, Proto A R, Abenavoli L M, et al. 2018. Spatial distribution of coarse root biomass and carbon in a high-density olive orchard: Effects of mechanical harvesting methods. Trees, 32: 919-931.
|
|
|
| [33] |
Uga Y, Sugimoto K, Ogawa S, et al. 2013. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nature Genetics, 45(9): 1097-1102.
|
|
|
| [34] |
van Noordwijk M, Spek L Y, de Willigen P. 1994. Proximal root diameter as predictor of total root size for fractal branching models: I. Theory. Plant and Soil, 164: 107-117.
|
|
|
| [35] |
van Noordwijk M, Mulia R. 2002. Functional branch analysis as tool for fractal scaling above- and belowground trees for their additive and non-additive properties. Ecological Modelling, 149(1-2): 41-51.
|
|
|
| [36] |
Wang G, Laga H, Jia J Y, et al. 2020. Statistical analysis and modeling of the geometry and topology of plant roots. Journal of Theoretical Biology, 486: 110108, doi: 10.1016/j.jtbi.2019.110108.
|
|
|
| [37] |
Yang X L, Zhang X M, Li Y L, et al. 2008. Analysis of root architecture and root adaptive strategy in the Taklimakan desert area of China. Chinese Journal of Plant Ecology, 32(6): 1268-1276. (in Chinese)
|
|
|
| [38] |
Zhao Y Y, Lu Z H, Xia J B, et al. 2015. Root architecture and adaptive strategy of 3 shrubs in Shell Bay in Yellow River Delta. Acta Ecologica Sinica, 35(6): 1688-1695. (in Chinese)
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|