| Research article |
|


|
|
|
| Effect of vegetation on soil bacteria and their potential functions for ecological restoration in the Hulun Buir Sandy Land, China |
YAN Ru1, FENG Wei1,2,*( ) ) |
1 Yanchi Research Station, School of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China
2 Engineering Research Center of Forestry Ecological Engineering of the Ministry of Education, Beijing Forestry University,Beijing 100083, China |
|
|
|
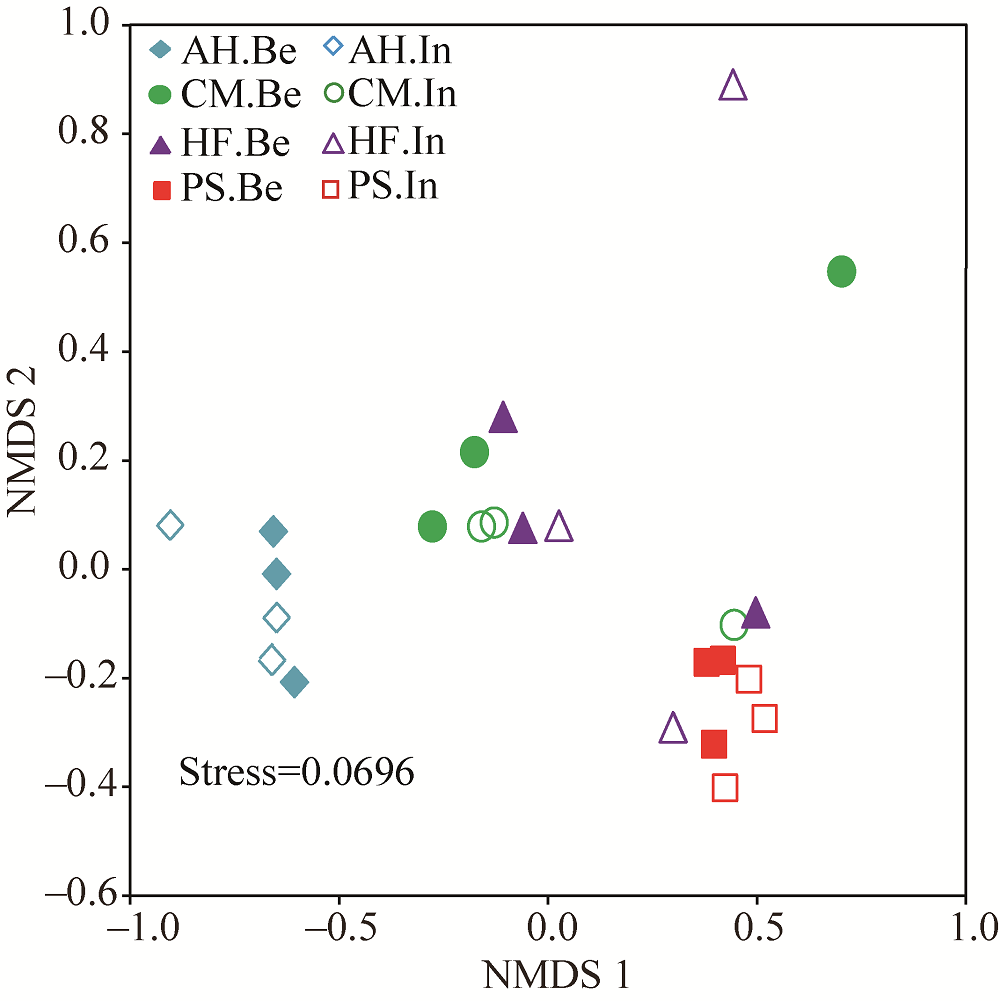
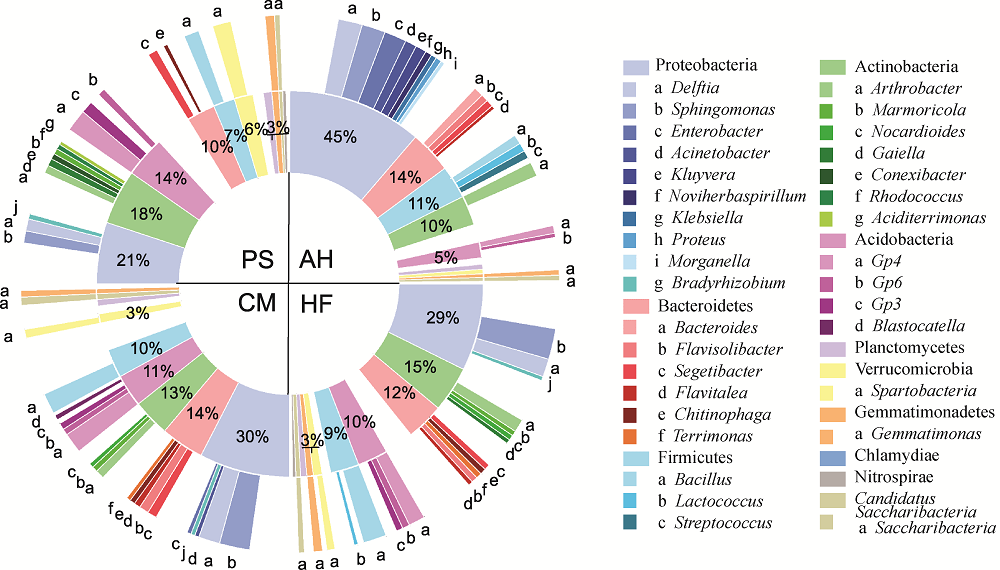
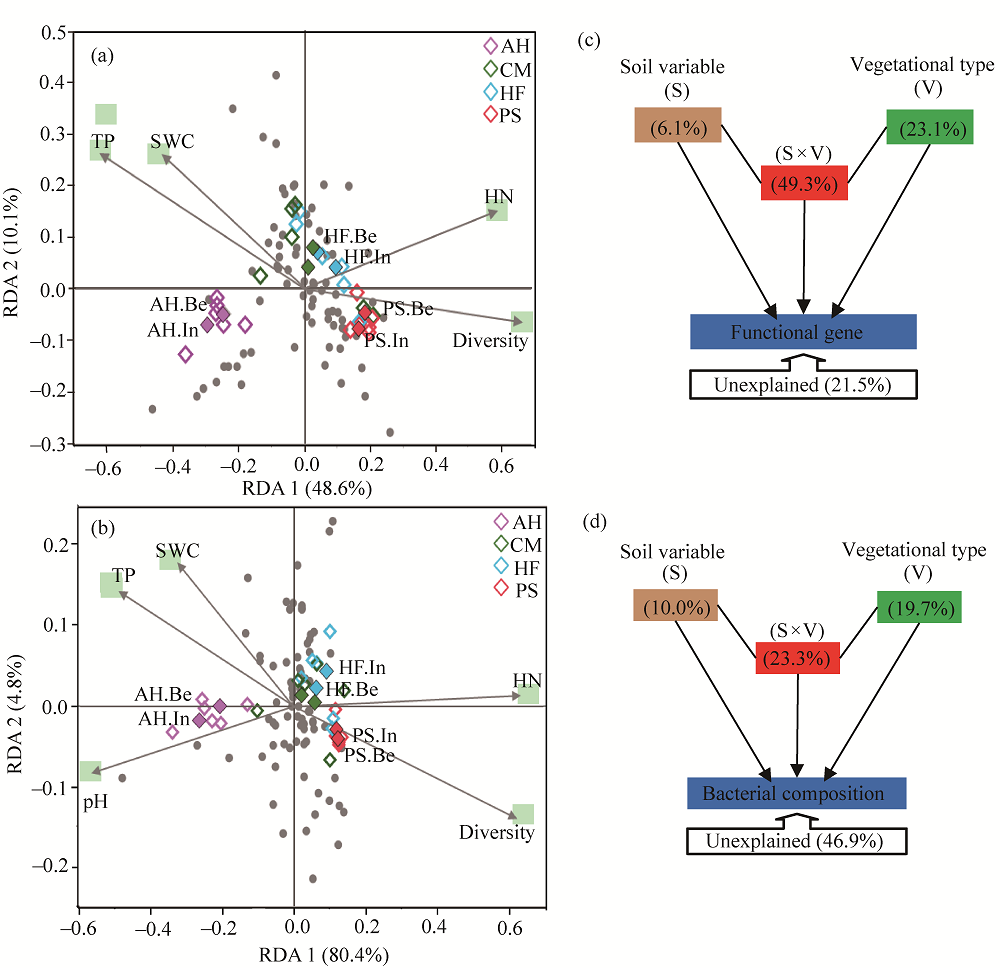
Abstract To date, much of research on revegetation has focused on soil microorganisms due to their contributions in the formation of soil and soil remediation process. However, little is known about the soil bacteria and their functions respond to the diverse vegetational types in the process of vegetation restoration. Effects of dominated vegetation, i.e., Artemisia halodendron Turcz Ex Bess, Caragana microphylla Lam., Hedysarum fruticosum Pall. and Pinus sylvestris L. on bacterial community structures and their potential functions in the Hulun Buir Sandy Land, China were determined using high-throughput 16S rRNA gene sequencing and phylogenetic investigation of communities by reconstruction of unobserved states (PICRUSt) in 2015. Although the dominant phyla of soil bacterial community among different types of vegetation, including Proteobacteria, Actinobacteria, Acidobacteria, Bacteroidetes and Firmicutes, were similar, the relative abundance of these dominant groups significantly differed, indicating that different types of vegetation might result in variations in the composition of soil bacterial community. In addition, functional genes of bacterial populations were similar among different types of vegetation, whereas its relative abundance was significantly differed. Most carbon fixation genes showed a high relative abundance in P. sylvestris, vs. recalcitrant carbon decomposition genes in A. halodendron, suggesting the variations in carbon cycling potential of different types of vegetation. Abundance of assimilatory nitrate reduction genes was the highest in P. sylvestris, vs. dissimilatory nitrate reduction and nitrate reductase genes in A. halodendron, indicating higher nitrogen gasification loss and lower nitrogen utilization gene functions in A. halodendron. The structures and functional genes of soil bacterial community showed marked sensitivities to different plant species, presenting the potentials for regulating soil carbon and nitrogen cycling.
|
|
Received: 06 September 2019
Published: 10 May 2020
|
|
Corresponding Authors:
|
| About author: *Corresponding author: FENG Wei (E-mail: weifeng@bjfu.edu.cn) |
|
|
| [1] |
Boeddinghaus R S, Marhan S, Berner Det al.2019. Plant functional trait shifts explain concurrent changes in the structure and function of grassland soil microbial communities. Journal of Ecology, 107(5): 2197-2210.
|
|
|
| [2] |
Cao H C, Chen R R, Wang L B, et al.2016. Soil pH, total phosphorus, climate and distance are the major factors influencing microbial activity at a regional spatial scale. Scientific Reports, 6: 25815.
|
|
|
| [3] |
Castro S P, Cleland E E, Wagner R, et al.2019. Soil microbial responses to drought and exotic plants shift carbon metabolism. The ISME Journal, 13: 1776-1787.
|
|
|
| [4] |
Che R X, Wang Y F, Li K X, et al.2019. Degraded patch formation significantly changed microbial community composition in alpine meadow soils. Soil and Tillage Research, 195: 104426.
|
|
|
| [5] |
Chen D M, Mi J, Chu P F, et al.2015. Patterns and drivers of soil microbial communities along a precipitation gradient on the Mongolian Plateau. Landscape Ecology, 30: 1669-1682.
|
|
|
| [6] |
Chen Y C, Sun J, Xie F T, et al.2015. Litter chemical structure is more important than species richness in affecting soil carbon and nitrogen dynamics including gas emissions from an alpine soil. Biology and Fertility of Soils, 51: 791-800.
|
|
|
| [7] |
Chen Y L, Xu T L, Veresoglou S D, et al.2017. Plant diversity represents the prevalent determinant of soil fungal community structure across temperate grasslands in northern China. Soil Biology and Biochemistry, 110: 12-21.
|
|
|
| [8] |
Cheng J M, Gang C C, Guo L, et al.2017. Modification in Grassland Ecology under the Influence of Changing Climatic and Land Use Conditions. Plant Ecology: Traditional Approaches to Recent Trends, 43.
|
|
|
| [9] |
Deng L, Peng C H, Huang C B, et al.2019. Drivers of soil microbial metabolic limitation changes along a vegetation restoration gradient on the Loess Plateau, China. Geoderma, 353: 188-200.
|
|
|
| [10] |
Du H S, Hasi E, Yang Y, et al.2012. Landscape pattern change and driving force of blowout distribution in the Hulun Buir Sandy Grassland. Sciences in Cold and Arid Regions, 4: 431-438.
|
|
|
| [11] |
Edgar R C.2010. Search and clustering orders of magnitude faster than BLAST. Bioinformatics, 26(19): 2460-2461.
|
|
|
| [12] |
Edgar R C, Haas B J, Clemente J C, et al.2011. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics, 27(16): 2194-2200.
|
|
|
| [13] |
Erisman J W, Sutton M A, Galloway J, et al.2008. How a century of ammonia synthesis changed the world. Nature Geoscience, 1: 636.
|
|
|
| [14] |
Evans S E, Wallenstein M D.2014. Climate change alters ecological strategies of soil bacteria. Ecology Letters, 17(2): 155-164.
|
|
|
| [15] |
Feng W, Zhang Y Q, Yan R, et al.2019. Dominant soil bacteria and their ecological attributes across the deserts in northern China. European Journal of Soil Science, 1-12. doi: https://doi.org/10.1111/ejss.12866.
|
|
|
| [16] |
Ferran G P, Virginia L, Yevgeniy M, et al.2013. Temperature drives the continental-scale distribution of key microbes in topsoil communities. Science, 340: 1574-1577.
|
|
|
| [17] |
Fierer N, Bradford M A, Jackson R B.2007. Toward an ecological classification of soil bacteria. Ecology, 88(6): 1354-1364.
|
|
|
| [18] |
Fierer N, Strickland M S, Liptzin D, et al.2009. Global patterns in belowground communities. Ecology Letters, 12(11): 1238-1249.
|
|
|
| [19] |
Fierer N, Ladau J, Clemente J C, et al.2013. Reconstructing the microbial diversity and function of pre-agricultural tallgrass prairie soils in the United States. Science, 342(6158): 621-624.
|
|
|
| [20] |
Fu B, Qi Y B, Chang Q R.2015. Impacts of revegetation management modes on soil properties and vegetation ecological restoration in degraded sandy grassland in farming-pastoral ecotone. International Journal of Agricultural and Biological Engineering, 8(1): 26-34.
|
|
|
| [21] |
Goberna M, Pascual J A, García C, et al.2007. Do plant clumps constitute microbial hotspots in semiarid Mediterranean patchy landscapes? Soil Biology and Biochemistry, 39(5): 1047-1054.
|
|
|
| [22] |
Goecks J, Nekrutenko A, Taylor J.2010. Galaxy: a comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biology, 11(8): R86.
|
|
|
| [23] |
Griffiths R I, Thomson B C, James P, et al.2011. The bacterial biogeography of British soils. Environmental Microbiology, 13(6): 1642-1654.
|
|
|
| [24] |
Gruber N, Galloway J N.2008. An Earth-system perspective of the global nitrogen cycle. Nature, 451: 293-296.
|
|
|
| [25] |
Guo J, Wang T, Xue X, et al.2010. Monitoring aeolian desertification process in Hulunbir grassland during 1975-2006, Northern China. Environmental Monitoring and Assessment, 166: 563-571.
|
|
|
| [26] |
Hacker N, Ebeling A, Gessler A, et al.2015. Plant diversity shapes microbe-rhizosphere effects on P mobilisation from organic matter in soil. Ecology Letters, 18(12): 1356-1365.
|
|
|
| [27] |
Hayat R, Ali S, Amara U, et al.2010. Soil beneficial bacteria and their role in plant growth promotion: a review. Annals of Microbiology, 60: 579-598.
|
|
|
| [28] |
Jangid K, Williams M A, Franzluebbers A J, et al.2011. Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biology and Biochemistry, 43(10): 2184-2193.
|
|
|
| [29] |
Jiang D M, Cao C Y, Zhang Y, et al.2014. Plantations of native shrub species restore soil microbial diversity in the Horqin Sandy Land, northeastern China. Journal of Arid Land, 6: 445-453.
|
|
|
| [30] |
Kim K Y, Jordan D, McDonald G.1998. Enterobacter agglomerans, phosphate solubilizing bacteria, and microbial activity in soil: effect of carbon sources. Soil Biology and Biochemistry, 30(8-9): 995-1003.
|
|
|
| [31] |
Kozich J J, Westcott S L, Baxter N T, et al.2013. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Applied and Environmental Microbiology, 79(17): 5112-5120.
|
|
|
| [32] |
Kuypers M M M, Marchant H K, Kartal B.2018. The microbial nitrogen-cycling network. Nature Reviews Microbiology, 16: 263-276.
|
|
|
| [33] |
Langille M G, Zaneveld J, Caporaso J G, et al.2013. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nature Biotechnology, 31: 814-821.
|
|
|
| [34] |
Larsbrink J, Tuveng T R, Pope P B, et al.2017. Proteomic insights into mannan degradation and protein secretion by the forest floor bacterium Chitinophaga pinensis. Journal of Proteomics, 156: 63-74.
|
|
|
| [35] |
Li J R, Okin G S, Alvarez L, et al.2007. Quantitative effects of vegetation cover on wind erosion and soil nutrient loss in a desert grassland of southern New Mexico, USA. Biogeochemistry, 85: 317-332.
|
|
|
| [36] |
Li X R, Zhang P, Su Y G, et al.2012. Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: A four-year field study. Catena, 97: 119-126.
|
|
|
| [37] |
Li Y F, Li Z W, Wang Z Y, et al.2017. Impacts of artificially planted vegetation on the ecological restoration of movable sand dunes in the Mugetan Desert, northeastern Qinghai-Tibet Plateau. International Journal of Sediment Research, 32: 277-287.
|
|
|
| [38] |
Liao C R, Liu B C, Xu Y N, et al.2019. Effect of topography and protecting barriers on revegetation of sandy land, Southern Tibetan Plateau. Scientific Reports, 9: 6501.
|
|
|
| [39] |
Liu Y M, Li X R, Xing Z S, et al.2013. Responses of soil microbial biomass and community composition to biological soil crusts in the revegetated areas of the Tengger Desert. Applied Soil Ecology, 65: 52-59.
|
|
|
| [40] |
Madsen E L.2011. Microorganisms and their roles in fundamental biogeochemical cycles. Currrent Opinion in Biotechnology, 22(3): 456-464.
|
|
|
| [41] |
Maestre F T, Quero J L, Gotelli N J, et al.2012. Plant species richness and ecosystem multifunctionality in global drylands. Science, 335(6065): 214-218.
|
|
|
| [42] |
Magoč T, Salzberg S L.2011. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics, 27(21): 2957-2963.
|
|
|
| [43] |
Marcel G, Richard D, Nico M.2008. The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters, 11(3): 296-310.
|
|
|
| [44] |
Miyasaka T, Okuro T, Miyamori E, et al.2014. Effects of different restoration measures and sand dune topography on short-and long-term vegetation restoration in northeast China. Journal of Arid Environments, 111: 1-6.
|
|
|
| [45] |
Neilson J W, Quade J, Ortiz M, et al.2012. Life at the hyperarid margin: novel bacterial diversity in arid soils of the Atacama Desert, Chile. Extremophiles, 16: 553-566.
|
|
|
| [46] |
Nelson M B, Martiny A C, Martiny J B.2016. Global biogeography of microbial nitrogen-cycling traits in soil. Proceedings of the National Academy of Sciences, 113(29): 8033-8040.
|
|
|
| [47] |
Orlando J, Alfaro M, Bravo L, et al.2010. Bacterial diversity and occurrence of ammonia-oxidizing bacteria in the Atacama Desert soil during a ''desert bloom'' event. Soil Biology and Biochemistry, 42(7): 1183-1188.
|
|
|
| [48] |
Özçelik M S, Gökbulak F, #x0015e;engönül K.2019. Effect of vegetation patch size on selected chemical properties of soils under semiarid climate conditions. Forestist, 69: 117-123.
|
|
|
| [49] |
Pei S F, Fu H, Wan C G.2008. Changes in soil properties and vegetation following exclosure and grazing in degraded Alxa desert steppe of Inner Mongolia, China. Agriculture, Ecosystems & Environment, 124(1-2): 33-39.
|
|
|
| [50] |
Prober S M, Leff J W, Bates S T, et al.2015. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecology Letters, 18(1): 85-95.
|
|
|
| [51] |
Schimel J P, Schaeffer S M.2012. Microbial control over carbon cycling in soil. Frontiers in Microbiology, 3: 348.
|
|
|
| [52] |
Schneider T, Keiblinger K M, Schmid E, et al.2012. Who is who in litter decomposition? Metaproteomics reveals major microbial players and their biogeochemical functions. The ISME Journal, 6: 1749-1762.
|
|
|
| [53] |
She W W, Bai Y X, Zhang Y Q, et al.2018. Resource availability drives responses of soil microbial communities to short-term precipitation and nitrogen addition in a desert shrubland. Frontiers in Microbiology, 9: 186.
|
|
|
| [54] |
Shihan A, Hättenschwiler S, Milcu A, et al.2017. Changes in soil microbial substrate utilization in response to altered litter diversity and precipitation in a Mediterranean shrubland. Biology and Fertility of Soils, 53: 171-185.
|
|
|
| [55] |
Sun L P, He LR, Wang G L, et al.2019. Natural vegetation restoration of Liaodong oak forests rapidly increased the content and ratio of inert carbon in soil macroaggregates. Journal of Arid Land, 11(6): 928-938.
|
|
|
| [56] |
Sun Y F, Zhang Y Q, Feng W, et al.2017. Effects of xeric shrubs on soil microbial communities in a desert in northern China. Plant and Soil, 414: 281-294.
|
|
|
| [57] |
Tiwari K, Gupta R K.2013. Diversity and isolation of rare actinomycetes: an overview. Critical Reviews in Microbiology, 39(3): 256-294.
|
|
|
| [58] |
Tu Q C, He Z L, Wu L Y, et al.2017. Metagenomic reconstruction of nitrogen cycling pathways in a CO2-enriched grassland ecosystem. Soil Biology and Biochemistry, 106: 99-108.
|
|
|
| [59] |
Unc A, Maggs-Kölling G, Marais E, et al.2019. Soil bacterial community associated with the dioecious Acanthosicyos horridus in the Namib Desert. Biology and Fertility of Soils, 55: 393-403.
|
|
|
| [60] |
Vered S T, Yosef S.2011. Soil microbial diversity in the vicinity of a Negev Desert shrub—Reaumuria negevensis. Microbial Ecology, 61: 64-81.
|
|
|
| [61] |
Voriskova J, Baldrian P.2013. Fungal community on decomposing leaf litter undergoes rapid successional changes. The ISME Journal, 7: 477-486.
|
|
|
| [62] |
Walkley A J, Black I A.1934. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science, 37: 29-38.
|
|
|
| [63] |
Wang B Z, Zhang C X, Liu J L, et al.2012. Microbial community changes along a land-use gradient of desert soil origin. Pedosphere, 22(5): 593-603.
doi: 10.1016/S1002-0160(12)60044-7
|
|
|
| [64] |
Wang S K, Zhao X Y, Zhang T H, et al.2013. Afforestation effects on soil microbial abundance, microbial biomass carbon and enzyme activity in dunes of Horqin Sandy Land, northeastern China. Sciences in Cold and Arid Regions, 5: 184-190.
|
|
|
| [65] |
Wang X B, van Nostrand J D, Deng Y, et al.2015. Scale-dependent effects of climate and geographic distance on bacterial diversity patterns across northern China's grasslands. FEMS Microbiology Ecology, 91(12): fiv133. doi: 10.1093/femsec/fiv133.
|
|
|
| [66] |
Wang X B, Lu X T, Yao J, et al.2017. Habitat-specific patterns and drivers of bacterial beta-diversity in China's drylands. The ISME Journal, 11: 1345-1358.
|
|
|
| [67] |
Wang X P, Quan G J, Pan Y X, et al.2013. Comparison of hydraulic behaviour of unvegetated and vegetation stabilized sand dunes in arid desert ecosystems. Ecohydrology, 6(2): 264-274.
|
|
|
| [68] |
Xiao E Z, Krumins V, Xiao T F, et al.2017. Depth-resolved microbial community analyses in two contrasting soil cores contaminated by antimony and arsenic. Environmental Pollution, 221: 244-255.
|
|
|
| [69] |
Yadav R S, Yadav B L, Chhipa B R, et al.2011. Soil biological properties under different tree based traditional agroforestry systems in a semi-arid region of Rajasthan, India. Agroforestry Systems, 81: 195-202.
|
|
|
| [70] |
Yang H T, Li X R, Wang Z R, et al.2014. Carbon sequestration capacity of shifting sand dune after establishing new vegetation in the Tengger Desert, northern China. Science of the Total Environment, 478: 1-11.
|
|
|
| [71] |
Yu L Z, Luo X S, Liu M, et al.2015. Diversity of ionizing radiation-resistant bacteria obtained from the Taklimakan Desert. Journal of Basic Microbiology, 55(1): 135-140.
|
|
|
| [72] |
Yuan J Y, Ouyang Z Y, Zheng H, et al.2012. Effects of different grassland restoration approaches on soil properties in the southeastern Horqin sandy land, northern China. Applied Soil Ecology, 61: 34-39. (in Chinese)
|
|
|
| [73] |
Zeng Q C, An S S, Liu Y.2017. Soil bacterial community response to vegetation succession after fencing in the grassland of China. Science of the Total Environment, 609: 2-10.
|
|
|
| [74] |
Zhang H F, Song X L, Wang C L, et al.2013a. The effects of different vegetation restoration patterns on soil bacterial diversity for sandy land in Hulunbeier. Acta Ecologica Sinica, 33(4): 211-216. (in Chinese)
|
|
|
| [75] |
Zhang H F, Li G, Song X L, et al.2013b. Changes in soil microbial functional diversity under different vegetation restoration patterns for Hulunbeier Sandy Land. Acta Ecologica Sinica, 33(1): 38-44. (in Chinese)
|
|
|
| [76] |
Zhang W, Zhang G S, Liu G X, et al.2012. Bacterial diversity and distribution in the southeast edge of the Tengger Desert and their correlation with soil enzyme activities. Journal of Environmental Sciences, 24: 2004-2011.
|
|
|
| [77] |
Zhang W, Xu Y D, Gao D X, et al.2019. Ecoenzymatic stoichiometry and nutrient dynamics along a revegetation chronosequence in the soils of abandoned land and Robinia pseudoacacia plantation on the Loess Plateau, China. Soil Biology and Biochemistry, 134: 1-14.
doi: 10.1016/j.soilbio.2019.03.017
|
|
|
| [78] |
Zhang X M, Zhang G M, Chen Q S, et al.2013. Soil bacterial communities respond to climate changes in a temperate steppe. PLoS ONE, 8: e78616.
|
|
|
| [79] |
Zhang Y, Cao C Y, Cui Z B, et al.2019. Soil bacterial community restoration along a chronosequence of sand-fixing plantations on moving sand dunes in the Horqin sandy land in northeast China. Journal of Arid Environments, 165: 81-87.
|
|
|
| [80] |
Zhao L N, Liu Y B, Wang Z R, et al.2019. Bacteria and fungi differentially contribute to carbon and nitrogen cycles during biological soil crust succession in arid ecosystems. Plant and Soil, 447: 379-392.
|
|
|
| [81] |
Zhou Y, Qin Y Q, Liu X D, et al.2019. Soil bacterial function associated with stylo (legume) and bahiagrass (grass) is affected more strongly by soil chemical property than by bacterial community composition. Frontiers in Microbiology, 10: 798.
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|