| Research article |
|


|
|
|
| Effects of water tables and nitrogen application on soil bacterial community diversity, network structure, and function in an alpine wetland, China |
HAN Yaoguang1,2,3,4, CHEN Kangyi2,5, SHEN Zhibo2,4, LI Keyi1,4, CHEN Mo1,4, HU Yang4,6, WANG Jiali2, JIA Hongtao2,4, ZHU Xinping3,4,7,*( ), YANG Zailei2,4 ), YANG Zailei2,4 |
1College of Grassland Science, Xinjiang Agricultural University, Urumqi 830052, China
2College of Resources and Environment, Xinjiang Agricultural University, Urumqi 830052, China
3College of Bioscience and Resources Environment, Beijing University of Agriculture, Beijing 102206, China
4Xinjiang Key Laboratory of Soil and Plant Ecological Processes, Urumqi 830052, China
5The Second Geological Brigade, Hebei Bureau of Geology and Mineral Exploration and Development, Tangshan 063000, China
6College of Natural Resources and Environment, Northwest A&F University, Yangling 712100, China
7Key Laboratory for North China Urban Agriculture of Ministry of Agriculture and Rural Affairs, Beijing 102206, China |
|
|
|
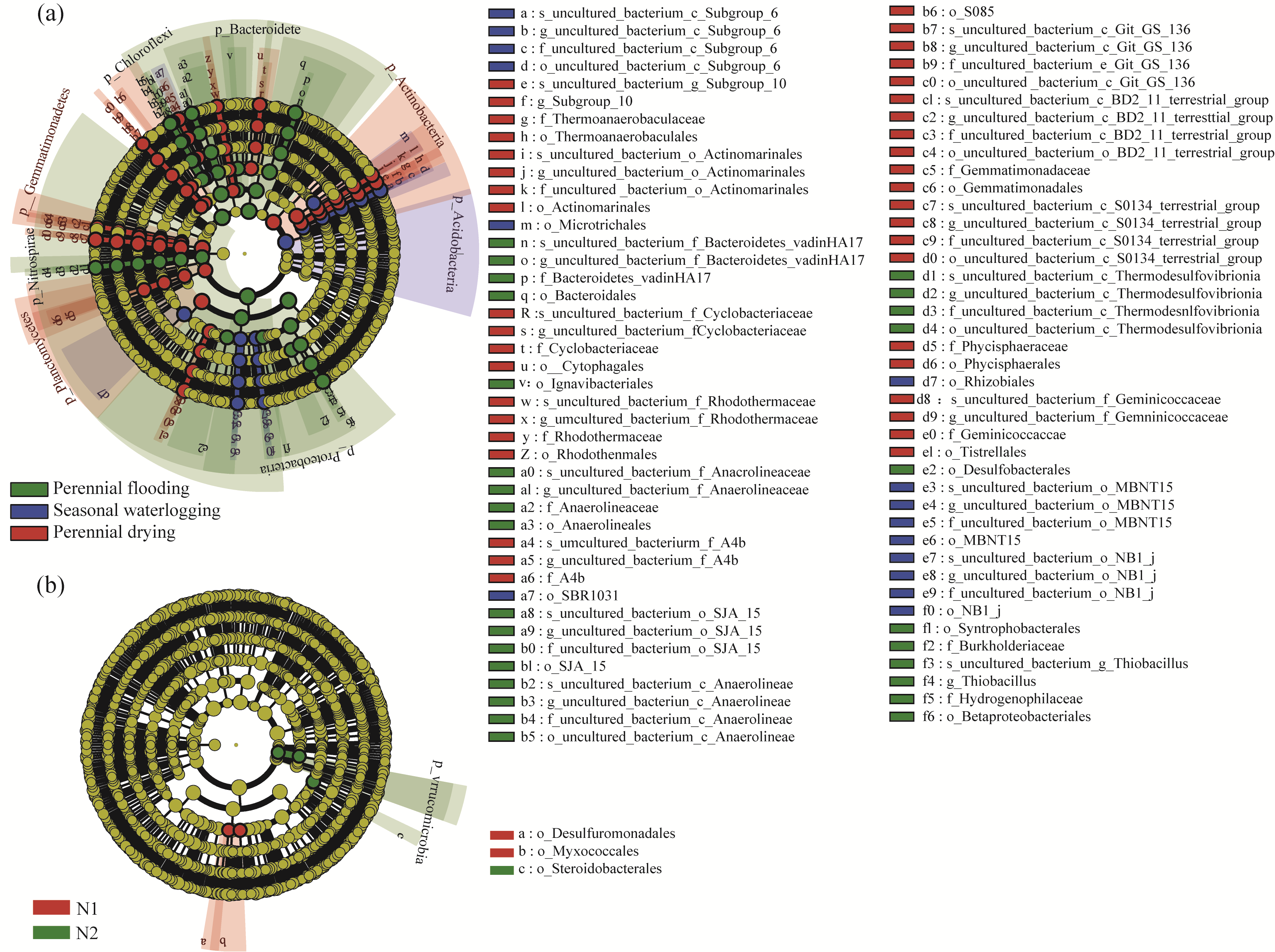
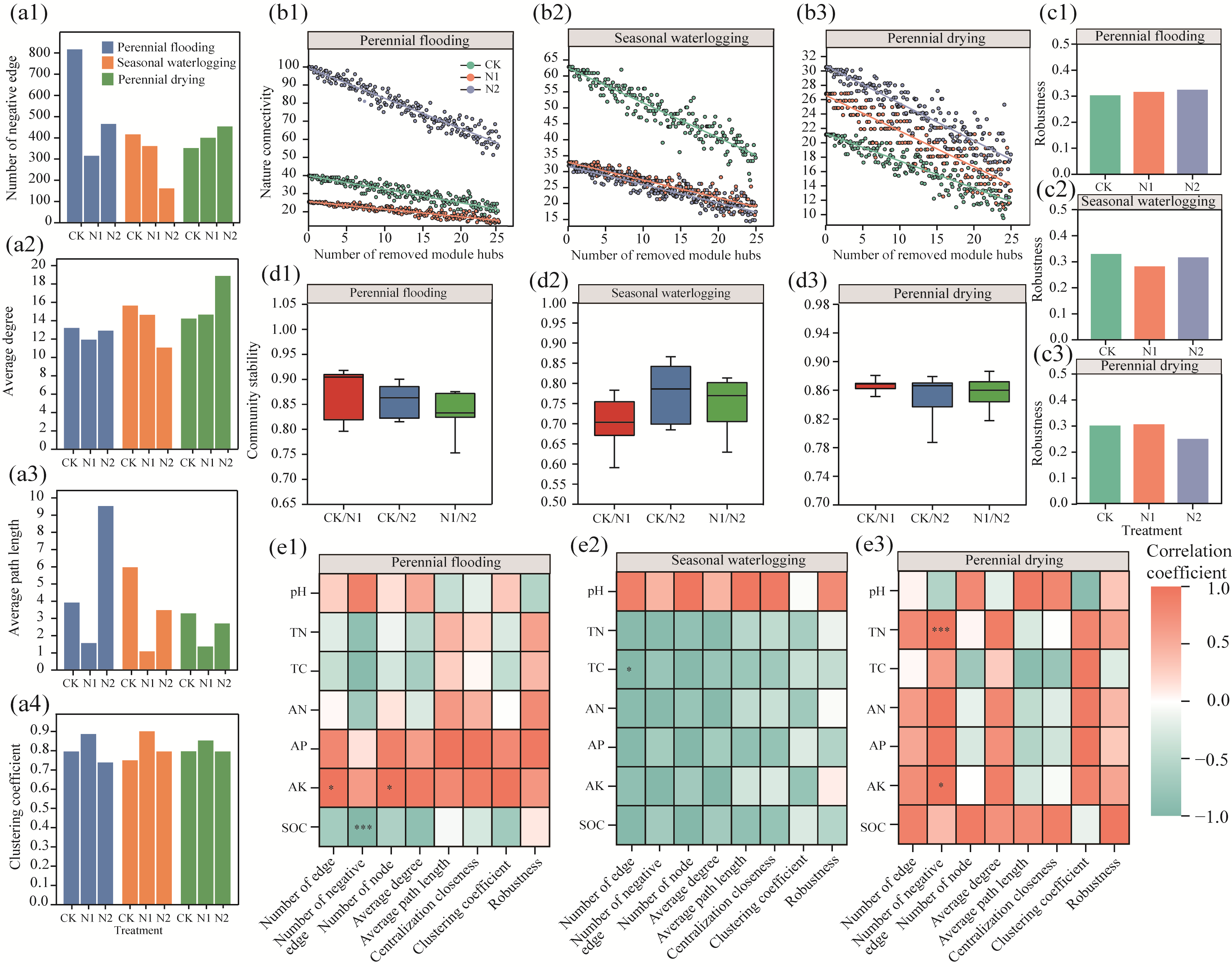
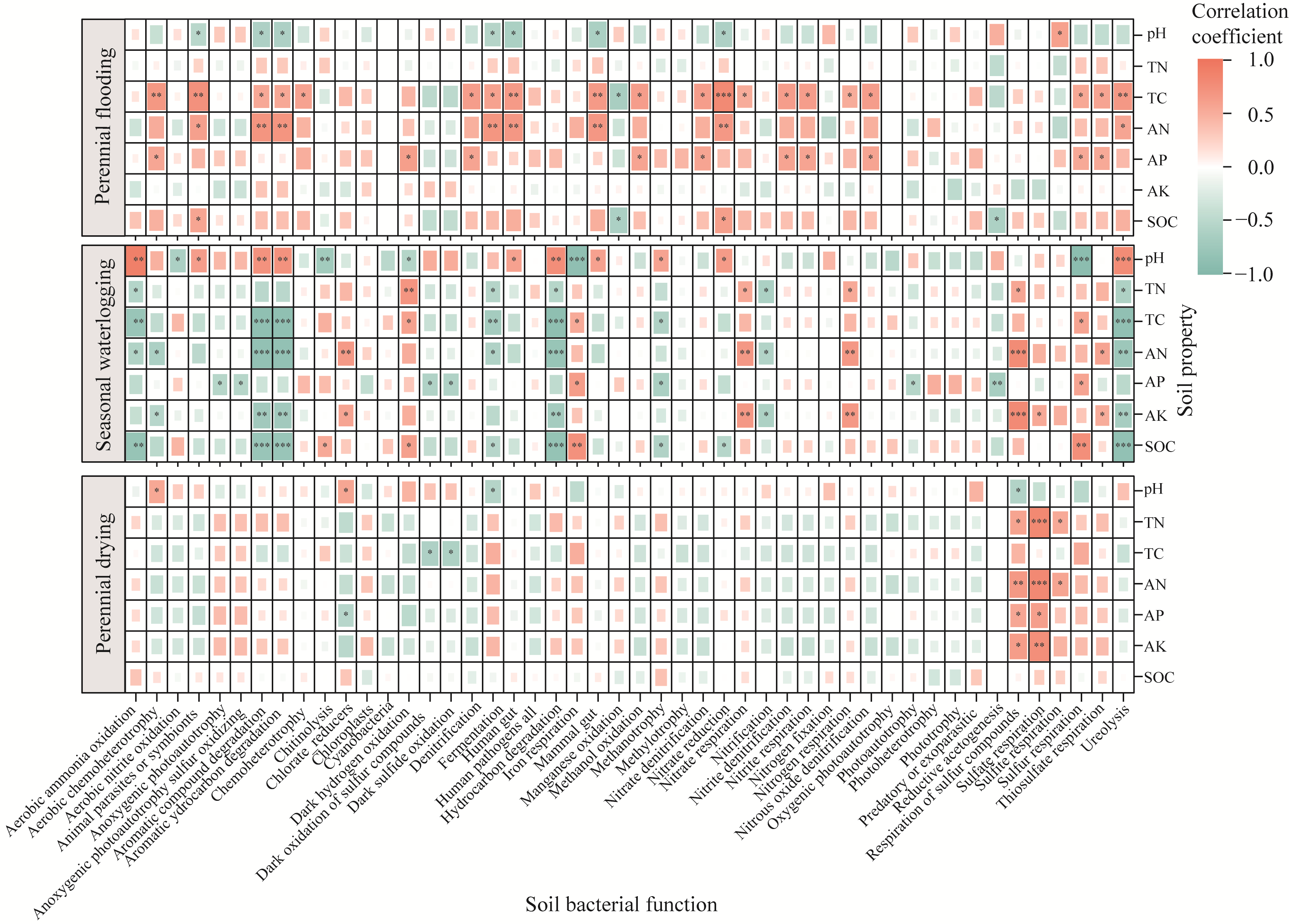
Abstract Nitrogen deposition and water tables are important factors to control soil microbial community structure. However, the specific effects and mechanisms of nitrogen deposition and water tables coupling on bacterial diversity, abundance, and community structure in arid alpine wetlands remain unclear. The nitrogen deposition (0, 10, and 20 kg N/(hm2•a)) experiments were conducted in the Bayinbulak alpine wetland with different water tables (perennial flooding, seasonal waterlogging, and perennial drying). The 16S rRNA (ribosomal ribonucleic acid) gene sequencing technology was employed to analyze the changes in bacterial community diversity, network structure, and function in the soil. Results indicated that bacterial diversity was the highest under seasonal waterlogging condition. However, nitrogen deposition only affected the bacterial Chao1 and beta diversity indices under seasonal waterlogging condition. The abundance of bacterial communities under different water tables showed significant differences at the phylum and genus levels. The dominant phylum, Proteobacteria, was sensitive to soil moisture and its abundance decreased with decreasing water tables. Although nitrogen deposition led to changes in bacterial abundance, such changes were small compared with the effects of water tables. Nitrogen deposition with 10 kg N/(hm2•a) decreased bacterial edge number, average path length, and robustness. However, perennial flooding and drying conditions could simply resist environmental changes caused by 20 kg N/(hm2•a) nitrogen deposition and their network structure remain unchanged. The sulfur cycle function was dominant under perennial flooding condition, and carbon and nitrogen cycle functions were dominant under seasonal waterlogging and perennial drying conditions. Nitrogen application increased the potential function of part of nitrogen cycle and decreased the potential function of sulfur cycle in bacterial community. In summary, composition of bacterial community in the arid alpine wetland was determined by water tables, and diversity of bacterial community was inhibited by a lower water table. Effect of nitrogen deposition on bacterial community structure and function depended on water tables.
|
|
Received: 24 July 2024
Published: 30 November 2024
|
|
Corresponding Authors:
*ZHU Xinping (E-mail: zhuxinping@bua.edu.cn)
|
| Cite this article:
HAN Yaoguang, CHEN Kangyi, SHEN Zhibo, LI Keyi, CHEN Mo, HU Yang, WANG Jiali, JIA Hongtao, ZHU Xinping, YANG Zailei. Effects of water tables and nitrogen application on soil bacterial community diversity, network structure, and function in an alpine wetland, China. Journal of Arid Land, 2024, 16(11): 1584-1603.
URL:
http://jal.xjegi.com/10.1007/s40333-024-0031-1 OR http://jal.xjegi.com/Y2024/V16/I11/1584
|
|
|
| [1] |
Abirami B, Radhakrishnan M, Kumaran S, et al. 2021. Impacts of global warming on marine microbial communities. Science of the Total Environment, 791: 147905, doi: 10.1016/j.scitotenv.2021.147905.
|
|
|
| [2] |
Abulaizi M, Chen M, Yang Z L, et al. 2023. Response of soil bacterial community to alpine wetland degradation in arid Central Asia. Frontiers in Plant Science, 13: 990597, doi: 10.3389/fpls.2022.990597.
|
|
|
| [3] |
Ackerman D, Millet D B, Chen X. 2019. Global estimates of inorganic nitrogen deposition across four decades. Global Biogeochemical Cycles, 33(1): 100-107.
doi: 10.1029/2018GB005990
|
|
|
| [4] |
Bao Y Y, Dolfing J, Guo Z Y, et al. 2021. Important ecophysiological roles of non-dominant Actinobacteria in plant residue decomposition, especially in less fertile soils. Microbiome, 9(1): 84, doi: 10.1186/s40168-021-01032-x.
|
|
|
| [5] |
Bobbink R, Hicks K, Galloway J, et al. 2010. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecological Applications, 20(1): 30-59.
pmid: 20349829
|
|
|
| [6] |
Che R X, Wang S P, Wang Y F, et al. 2019. Total and active soil fungal community profiles were significantly altered by six years of warming but not by grazing. Soil Biology and Biochemistry, 139: 107611, doi: 10.1016/j.soilbio.2019.107611.
|
|
|
| [7] |
Chen M, Zhu X P, Zhao C Y, et al. 2021. Rapid microbial community evolution in initial Carex litter decomposition stages in Bayinbuluk alpine wetland during the freeze-thaw period. Ecological Indicators, 121: 107180, doi: 10.1016/j.ecolind.2020.107180.
|
|
|
| [8] |
Chen W J, Zhou H K, Wu Y, et al. 2020. Direct and indirect influences of long-term fertilization on microbial carbon and nitrogen cycles in an alpine grassland. Soil Biology and Biochemistry, 149: 107922, doi: 10.1016/j.soilbio.2020.107922.
|
|
|
| [9] |
Chowdhury R T, Lee J Y, Bottos E M, et al. 2019. Metaphenomic responses of a native prairie soil microbiome to moisture perturbations. mSystems, 4(4): e00061-19, doi: 10.1128/MsYSTEMS.00061-19.
|
|
|
| [10] |
Dai Z M, Su W Q, Chen H H, et al. 2018. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of Actinobacteria and Proteobacteria in agro-ecosystems across the globe. Global Change Biology, 24: 3452-3461.
doi: 10.1111/gcb.14163
pmid: 29645398
|
|
|
| [11] |
de Vries F T, Griffiths R I, Bailey M, et al. 2018. Soil bacterial networks are less stable under drought than fungal networks. Nature Communications, 9(1): 3033, doi: 10.1038/s41467-018-05516-7.
|
|
|
| [12] |
Deng Y, Che R, Wu Y, et al. 2015. A review of the physiological and ecological characteristics of methanotrophs and methanotrophic community diversity in the natural wetlands. Acta Ecologica Sinica, 35(14): 4579-4591. (in Chinese)
|
|
|
| [13] |
Fan S, Qin J, Sun H, et al. 2022. Alpine soil microbial community structure and diversity are largely influenced by moisture content in the Zoige Wetland. International Journal of Environmental Science and Technology, 19(5): 4369-4378.
|
|
|
| [14] |
Götzenberger L, de Bello F, Bråthen K A, et al. 2012. Ecological assembly rules in plant communities-approaches, patterns and prospects. Biological Reviews, 87(1): 111-127.
|
|
|
| [15] |
Griffiths R, Whiteley A, O'donnell A, et al. 2003. Physiological and community responses of established grassland bacterial populations to water stress. Applied and Environmental Microbiology, 69(12): 6961-6968.
doi: 10.1128/AEM.69.12.6961-6968.2003
pmid: 14660337
|
|
|
| [16] |
Hoek T, Axelrod K, Biancalani T, et al. 2016. Resource availability modulates the cooperative and competitive nature of a microbial cross-feeding mutualism. PLoS Biology, 14(8): e1002540, doi: 10.1371/journal.pbio.1002540.
|
|
|
| [17] |
Hu Y, Chen M, Yang Z L, et al. 2021. Soil microbial community response to nitrogen application on a swamp meadow in the arid region of Central Asia. Front Microbiol, 12: 797306, doi: 10.3389/fmicb.2021.797306.
|
|
|
| [18] |
Hu Y, Cong M F, Yan H, et al. 2023. Effects of biochar addition on aeolian soil microbial community assembly and structure. Applied Microbiology and Biotechnology, 107(11): 3829-3845.
|
|
|
| [19] |
Huang L B, Zhang G L, Bai J H, et al. 2020. Desalinization via freshwater restoration highly improved microbial diversity, co-occurrence patterns and functions in coastal wetland soils. Science of the Total Environment, 765: 142769, doi: 10.1016/j.scitotenv.2020.142769.
|
|
|
| [20] |
Isobe K, Allison S D, Khalili B, et al. 2019. Phylogenetic conservation of bacterial responses to soil nitrogen addition across continents. Nature Communications, 10(1): 2499, doi: 10.1038/s41467-019-10390-y.
|
|
|
| [21] |
Jassey V E J, Reczuga M K, Zielińska M, et al. 2018. Tipping point in plant-fungal interactions under severe drought causes abrupt rise in peatland ecosystem respiration. Global Change Biology, 24(3): 972-986.
doi: 10.1111/gcb.13928
pmid: 28991408
|
|
|
| [22] |
Jiao S, Qi J J, Jin C J, et al. 2022. Core phylotypes enhance the resistance of soil microbiome to environmental changes to maintain multifunctionality in agricultural ecosystems. Global Change Biology, 28(22): 6653-6664.
|
|
|
| [23] |
Kim S Y, Lee S H, Freeman C, et al. 2008. Comparative analysis of soil microbial communities and their responses to the short-term drought in bog, fen, and riparian wetlands. Soil Biology and Biochemistry, 40(11): 2874-2880.
|
|
|
| [24] |
Li K H, Liu X J, Song L, et al. 2015. Response of alpine grassland to elevated nitrogen deposition and water supply in China. Oecologia, 177(1): 65-72.
doi: 10.1007/s00442-014-3122-4
pmid: 25407621
|
|
|
| [25] |
Li Q, Xiao R, Sun L H, et al. 2017a. Bacterial and diazotrophic diversities of endophytes in Dendrobium catenatum determined through barcoded pyrosequencing. PLoS ONE, 12(9): e0184717, doi: 10.1371/journal.pone.0184717.
|
|
|
| [26] |
Li M, Yu X W, Kang H, et al. 2017b. Concentrations and size distributions of bacteria-containing particles over oceans from China to the Arctic Ocean. Atmosphere, 8: 82, doi: 10.3390/atmos8050082.
|
|
|
| [27] |
Li K H, Liu X J, Geng F Z, et al. 2021a. Inorganic nitrogen deposition in arid land ecosystems of Central Asia. Environmental Science and Pollution Research, 28(24): 31861-31871.
|
|
|
| [28] |
Li M C, Wei G S, Liu J A, et al. 2022. Effects of nitrate exposure on nitrate reduction processes in the wetland sediments from the Yellow River estuary. Estuaries and Coasts, 45(2): 315-330.
|
|
|
| [29] |
Li W C, Sheng H Y, Ekawati D, et al. 2019. Variations in the compositions of soil bacterial and fungal communities due to microhabitat effects induced by simulated nitrogen deposition of a bamboo forest in wetland. Forests, 10(12): 1098, doi: 10.3390/f10121098.
|
|
|
| [30] |
Li Y T, He J S, Wang H, et al. 2021b. Lowered water table causes species substitution while nitrogen amendment causes species loss in alpine wetland microbial communities. Pedosphere, 31(6): 912-922.
|
|
|
| [31] |
Liu D D, Ju W L, Jin X L, et al. 2021. Associated soil aggregate nutrients and controlling factors on aggregate stability in semiarid grassland under different grazing prohibition timeframes. Science of the Total Environment, 777: 146104, doi: 10.1016/j.scitotenv.2021.146104.
|
|
|
| [32] |
Liu W B, Ling N, Guo J J, et al. 2020a. Legacy effects of 8-year nitrogen inputs on bacterial assemblage in wheat rhizosphere. Biology and Fertility of Soils, 56(5): 583-596.
|
|
|
| [33] |
Liu X J, Duan L, Mo J M, et al. 2011. Nitrogen deposition and its ecological impact in China: An overview. Environmental Pollution, 159(10): 2251-2264.
doi: 10.1016/j.envpol.2010.08.002
pmid: 20828899
|
|
|
| [34] |
Liu X J, Zhang Y, Han W H, et al. 2013. Enhanced nitrogen deposition over China. Nature, 494: 459-462.
|
|
|
| [35] |
Liu X Y, Hu S H, Sun R, et al. 2021. Dissolved oxygen disturbs nitrate transformation by modifying microbial community, co-occurrence networks, and functional genes during aerobic-anoxic transition. Science of the Total Environment, 790: 148245, doi: 10.1016/j.scitotenv.2021.148245.
|
|
|
| [36] |
Liu Y, Ren Z, Qu X D, et al. 2020b. Microbial community structure and functional properties in permanently and seasonally flooded areas in Poyang Lake. Scientific Reports, 10(1): 4819, doi: 10.1038/s41598-020-61569-z.
|
|
|
| [37] |
Liusui Y H, Zhu X P, Li D P, et al. 2019. Soil aggregate and intra-aggregate carbon fractions associated with vegetation succession in an alpine wetland of Northwest China. Catena, 181: 104107, doi: 10.1016/j.catena.2019.104107.
|
|
|
| [38] |
Lu G R, Xie B H, Cagle G, et al. 2021. Effects of simulated nitrogen deposition on soil microbial community diversity in coastal wetland of the Yellow River Delta. Science of the Total Environment, 757: 143825, doi: 10.1016/j.scitotenv.2020.143825.
|
|
|
| [39] |
Ma X Y, Song Y Y, Song C C, et al. 2021. Effect of Nitrogen addition on soil microbial functional gene abundance and community diversity in permafrost peatland. Microorganisms, 9(12): 9122498, doi: 10.3390/microorganisms9122498.
|
|
|
| [40] |
Ma X Y, Wang T X, Shi Z, et al. 2022. Long-term nitrogen deposition enhances microbial capacities in soil carbon stabilization but reduces network complexity. Microbiome, 10(1): 112, doi: 10.1186/s40168-022-01309-9.
|
|
|
| [41] |
Nicolás C, Martin-Bertelsen T, Floudas D, et al. 2019. The soil organic matter decomposition mechanisms in ectomycorrhizal fungi are tuned for liberating soil organic nitrogen. Multidisciplinary Journal of Microbial Ecology, 13(4): 977-988.
|
|
|
| [42] |
Peralta A L, Ludmer S, Kent A D. 2013. Hydrologic history influences microbial community composition and nitrogen cycling under experimental drying/wetting treatments. Soil Biology and Biochemistry, 66: 29-37.
|
|
|
| [43] |
Potter C, Freeman C, Golyshin P N, et al. 2017. Subtle shifts in microbial communities occur alongside the release of carbon induced by drought and rewetting in contrasting peatland ecosystems. Scientific Reports, 7: 11314, doi: 10.1038/s41598-017-11546-w.
pmid: 28900257
|
|
|
| [44] |
Quast C, Pruesse E, Yilma P, et al. 2013. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research, 41: 590-596.
|
|
|
| [45] |
Ramirez K S, Lauber C L, Knight R, et al. 2010. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology, 91(12): 3463-3470.
pmid: 21302816
|
|
|
| [46] |
Reed D W, Smith J M, Francis C A, et al. 2010. Responses of ammonia-oxidizing bacterial and archaeal populations to organic nitrogen amendments in low-nutrient groundwater. Applied and Environmental Microbiology, 76(8): 2517-2523.
doi: 10.1128/AEM.02436-09
pmid: 20190081
|
|
|
| [47] |
RoyChowdhury T, Bramer L M, Brown J, et al. 2022. Soil metabolomics predict microbial taxa as biomarkers of moisture status in soils from a Tidal wetland. Microorganisms, 10(8): 1653, doi: 10.3390/microorganisms10081653.
|
|
|
| [48] |
Shi X Z, Hu H W, Wang J Q, et al. 2018. Niche separation of Comammox nitrospira and canonical ammonia oxidizers in an acidic subtropical forest soil under long-term nitrogen deposition. Soil Biology and Biochemistry, 126: 114-122.
|
|
|
| [49] |
Siebielec S, Siebielec G, Klimkowicz-Pawlas A, et al. 2020. Impact of water stress on microbial community and activity in sandy and loamy soils. Agronomy, 10(9): 1429, doi: 10.3390/agronomy10091429.
|
|
|
| [50] |
Sokol N W, Slessarev E, Marschmann G L, et al. 2022. Life and death in the soil microbiome: How ecological processes influence biogeochemistry. Nature Reviews Microbiology, 20(7): 415-430.
doi: 10.1038/s41579-022-00695-z
pmid: 35228712
|
|
|
| [51] |
Stevens C J, Basto S, Bell M D, et al. 2022. Research progress on the impact of nitrogen deposition on global grasslands. Frontiers of Agricultural Science and Engineering, 9(3): 425-444.
doi: 10.15302/J-FASE-2022457
|
|
|
| [52] |
Sun J B, Li W B, Li C Q, et al. 2020. Effect of different rates of nitrogen fertilization on crop yield, soil properties and leaf physiological attributes in banana under subtropical regions of China. Frontiers in Plant Science, 11: 613760, doi: 10.3389/fpls.2020.613760.
|
|
|
| [53] |
Sun M Y, Li M C, Zhou Y Q, et al. 2023. Nitrogen deposition enhances the deterministic process of the prokaryotic community and increases the complexity of the microbial co-network in coastal wetlands. Science of the Total Environment, 856(1): 158939, doi: 10.1016/j.scitotenv.2022.158939.
|
|
|
| [54] |
Tian G L, Qiu H S, Li D W, et al. 2022. Little environmental adaptation and high stability of bacterial communities in rhizosphere rather than bulk soils in rice fields. Applied Soil Ecology, 169: 104183, doi: 10.1016/j.apsoil.2021.104183.
|
|
|
| [55] |
Treseder K K. 2008. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecology Letter, 11(10): 1111-1120.
|
|
|
| [56] |
Uhlířová E, Elhottová D, Tříska J, et al. 2008. Physiology and microbial community structure in soil at extreme water content. Folia Microbiologica, 50(2): 161-166.
|
|
|
| [57] |
Urbanová Z, Bárta J. 2016. Effects of long-term drainage on microbial community composition vary between peatland types. Soil Biology and Biochemistry, 92:16-26.
|
|
|
| [58] |
Wang M Y, Weng X H, Zhang R T, et al. 2022a. The diversity and composition of soil microbial community differ in three typical wetland types of the Sanjiang Plain, Northeastern China. Sustainability, 14(21): 14394, doi: 10.3390/su142114394.
|
|
|
| [59] |
Wang Q F, Ma M C, Jiang X, et al. 2019. Impact of 36 years of nitrogen fertilization on microbial community composition and soil carbon cycling-related enzyme activities in rhizospheres and bulk soils in northeast China. Applied Soil Ecology, 136: 148-157.
|
|
|
| [60] |
Wang X D, Feng J G, Ao G, et al. 2023. Globally nitrogen addition alters soil microbial community structure, but has minor effects on soil microbial diversity and richness. Soil Biology and Biochemistry, 179: 108982, doi: 10.1016/j.soilbio.2023.108982.
|
|
|
| [61] |
Wang Y T, Xie Y Z, Ma H B, et al. 2022b. Responses of soil microbial communities and networks to precipitation change in a typical steppe ecosystem of the Loess Plateau. Microorganisms, 10(4): 817, doi: 10.3390/microorganisms10040817.
|
|
|
| [62] |
Wen T, Xie P H, Yang S D, et al. 2022. ggClusterNet: An R package for microbiome network analysis and modularity-based multiple network layouts. iMeta, 1(3): e32, doi: 10.1002/imt2.32.
|
|
|
| [63] |
Williams M A, Rice C W. 2007. Seven years of enhanced water availability influences the physiological, structural, and functional attributes of a soil microbial community. Applied Soil Ecology, 35(3): 535-545.
|
|
|
| [64] |
Yan B S, Sun L P, Li J J, et al. 2020a. Change in composition and potential functional genes of soil bacterial and fungal communities with secondary succession in Quercus liaotwigensis forests of the Loess Plateau, western China. Geoderma, 364: 114199, doi: 10.1016/j.geoderma.2020.114199.
|
|
|
| [65] |
Yan G Y, Xing Y J, Han S J, et al. 2020b. Long-time precipitation reduction and nitrogen deposition increase alter soil nitrogen dynamic by influencing soil bacterial communities and functional groups. Pedosphere, 30(3): 363-377.
|
|
|
| [66] |
Yang Y, Chen X L, Liu L X, et al. 2022. Nitrogen fertilization weakens the linkage between soil carbon and microbial diversity: A global meta-analysis. Global Change Biology, 28(21): 6446-6461.
|
|
|
| [67] |
Yin Z Y, Yu X F, Zou Y C, et al. 2022. Nitrogen addition effects on wetland soils depend on environmental factors and nitrogen addition methods: A meta-analysis. Water, 14(11): 1748, doi: 10.3390/w14111748.
|
|
|
| [68] |
Yu S, Ehrenfeld J G. 2009. The effects of changes in soil moisture on nitrogen cycling in acid wetland types of the New Jersey Pinelands (USA). Soil Biology and Biochemistry, 41(12): 2394-2405.
|
|
|
| [69] |
Yu X L, Hu R W, Tao M, et al. 2023. Microbially driven sulfur cycling in the River-Wetland-Ocean continuum. Ocean-Land- Atmosphere Research, 2: 0027, doi: 10.34133/olar.0027.
|
|
|
| [70] |
Yuan J Y, Liang Y L, Zhuo M C, et al. 2023. Soil nitrogen and carbon storages and carbon pool management index under sustainable conservation tillage strategy. Frontiers in Ecology and Evolution, 10: 1082624, doi: 10.3389/fevo.2022.1082624.
|
|
|
| [71] |
Yuan M M, Guo X, Wu L W, et al. 2021. Climate warming enhances microbial network complexity and stability. Nature Climate Change, 11(4): 343-348.
|
|
|
| [72] |
Zeng J, Liu X J, Song L, et al. 2016. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biology and Biochemistry, 92: 41-49.
|
|
|
| [73] |
Zhang B, Niu Z G, Zhang D Q, et al. 2022a. Dynamic changes and driving forces of alpine wetlands on the Qinghai-Tibetan plateau based on long-term time series satellite data: A case study in the Gansu Maqu wetlands. Remote Sensing, 14(17): 4147, doi: 10.3390/rs14174147.
|
|
|
| [74] |
Zhang W T, Kang X M, Kang E Z, et al. 2022b. Soil water content, carbon, and nitrogen determine the abundances of methanogens, methanotrophs, and methane emission in the Zoige alpine wetland. Journal of Soils and Sediments, 22(2): 470-481.
|
|
|
| [75] |
Zhang Y M, Naafs B D A, Huang X Y, et al. 2022c. Variations in wetland hydrology drive rapid changes in the microbial community, carbon metabolic activity, and greenhouse gas fluxes. Geochimica et Cosmochimica Acta, 317: 269-285.
|
|
|
| [76] |
Zhang Z H, Li T, Shao P S, et al. 2023. Effects of short-term nitrogen addition on rhizosphere and bulk soil bacterial community structure of three halophytes in the Yellow River Delta. Land Degradation & Development, 34(11): 3281-3294.
|
|
|
| [77] |
Zhou H, Zhang D G, Jiang Z H, et al. 2019. Changes in the soil microbial communities of alpine steppe at Qinghai-Tibetan Plateau under different degradation levels. Science of the Total Environment, 651: 2281-2291.
|
|
|
| [78] |
Zhu X M, Zhang W, Chen H, et al. 2015. Impacts of nitrogen deposition on soil nitrogen cycle in forest ecosystems: A review. Acta Ecologica Sinica, 35(3): 35-43. (in Chinese)
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|