| Research article |
|

|
|
|
| Hydro-saline synergy regulates ecosystem multifunctionality via microbial biomass in semi-arid grasslands, China |
HU Jinpeng, HE Yuanyuan, LI Yuanhong, ZHANG Yuewei, ZHANG Jinlin*( ) ) |
| State Key Laboratory of Herbage Improvement and Grassland Agro-ecosystems, Engineering Research Center of Grassland Industry, Ministry of Education; Center for Grassland Microbiome; College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou, 730000, China |
|
|
|
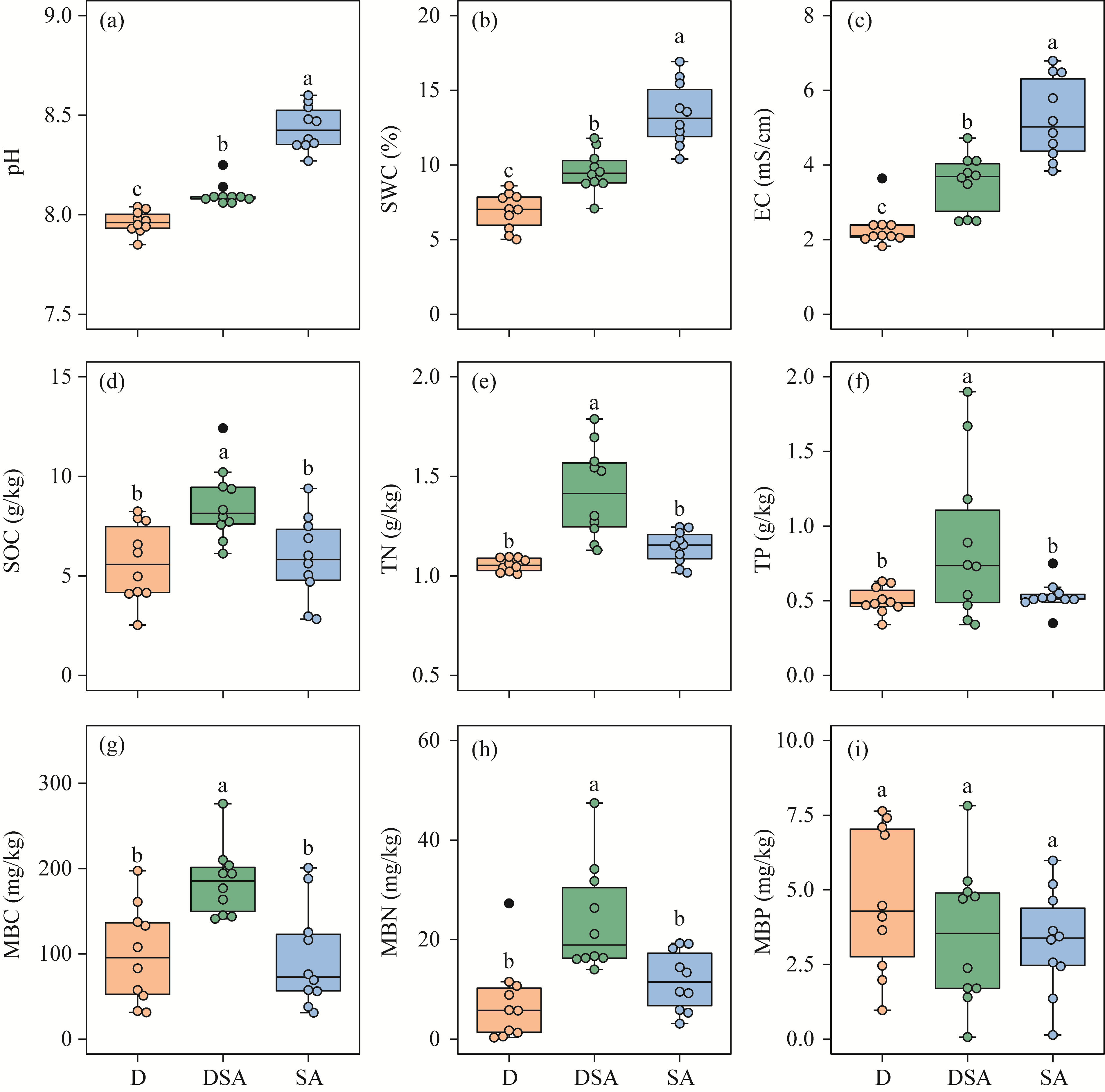
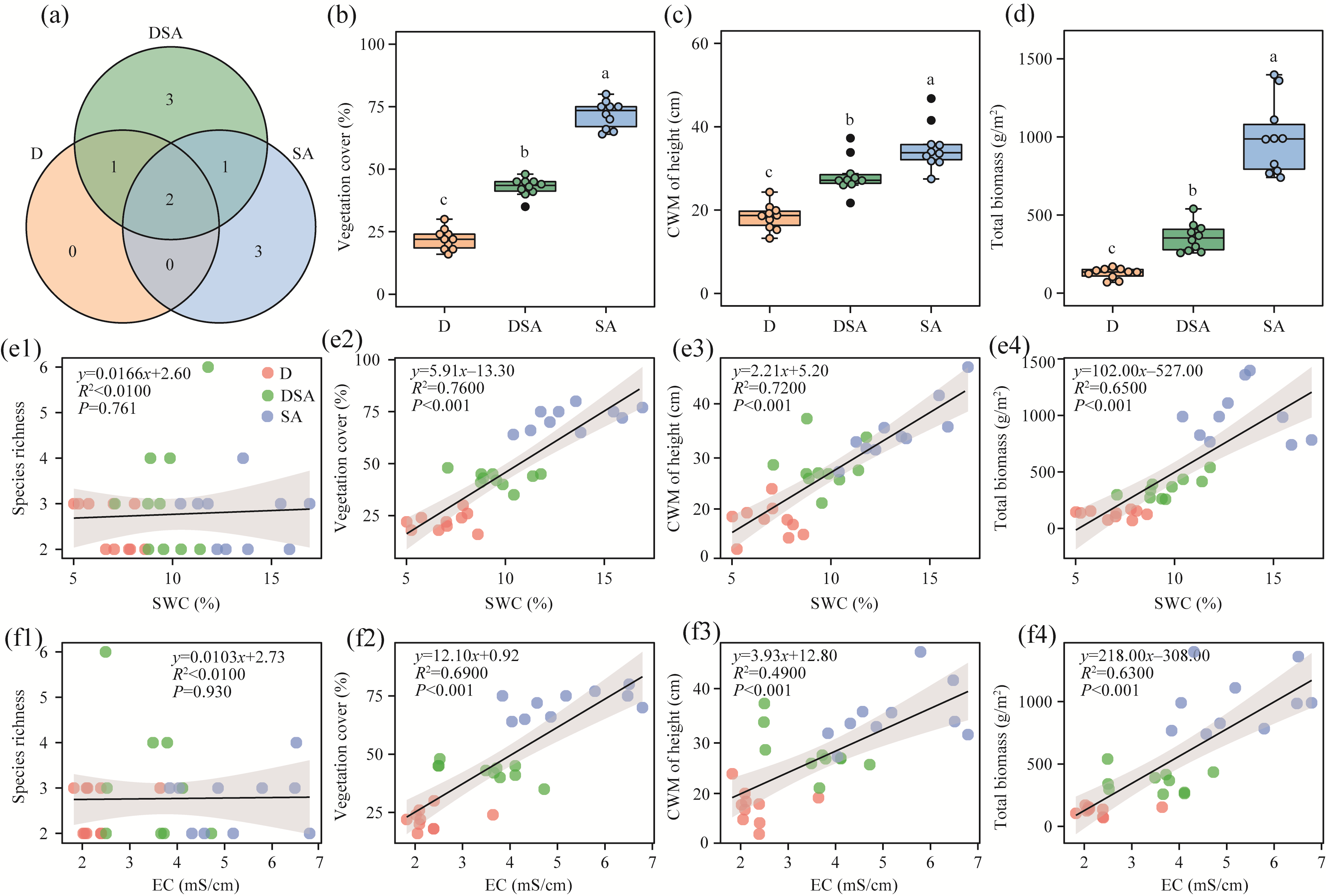
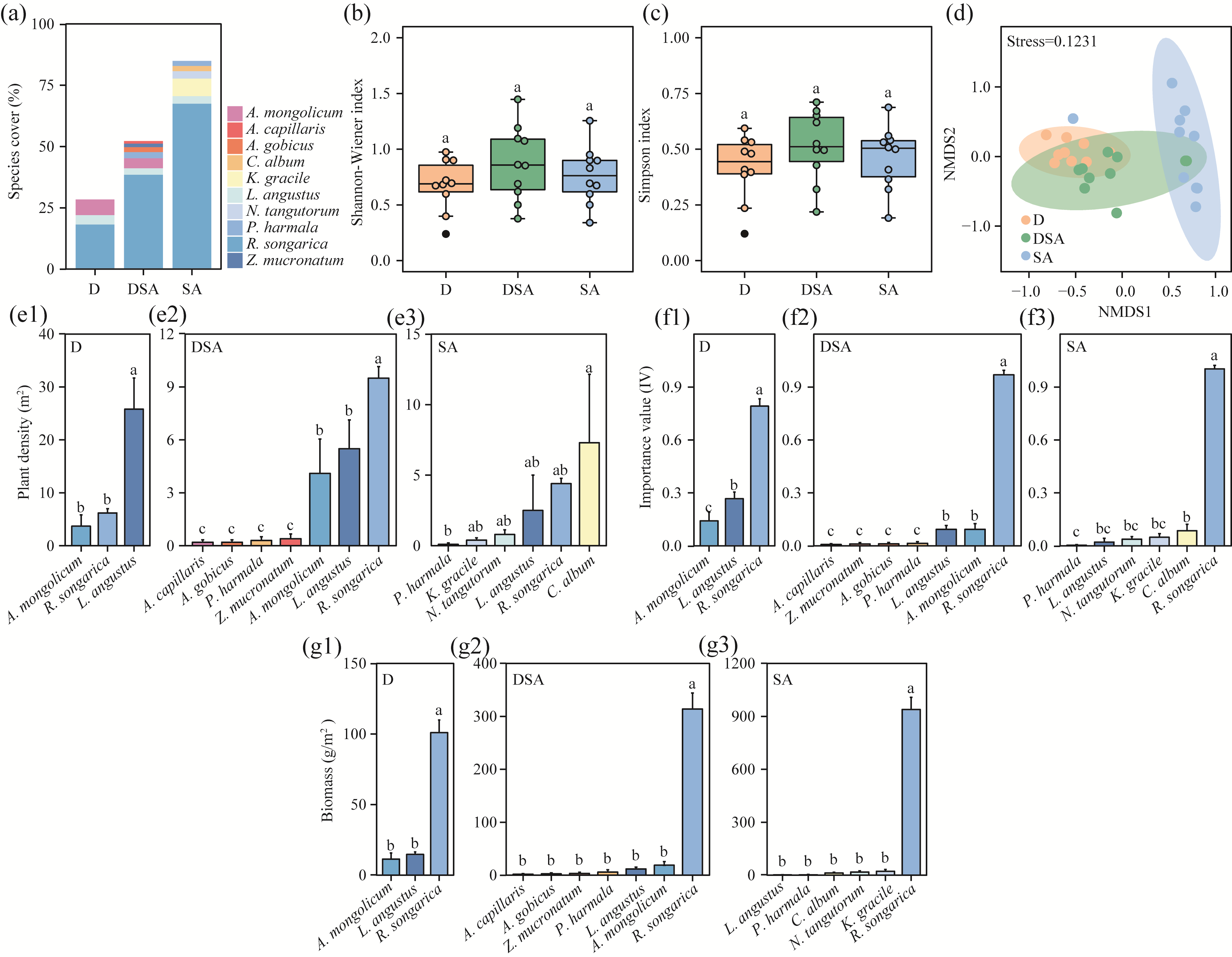
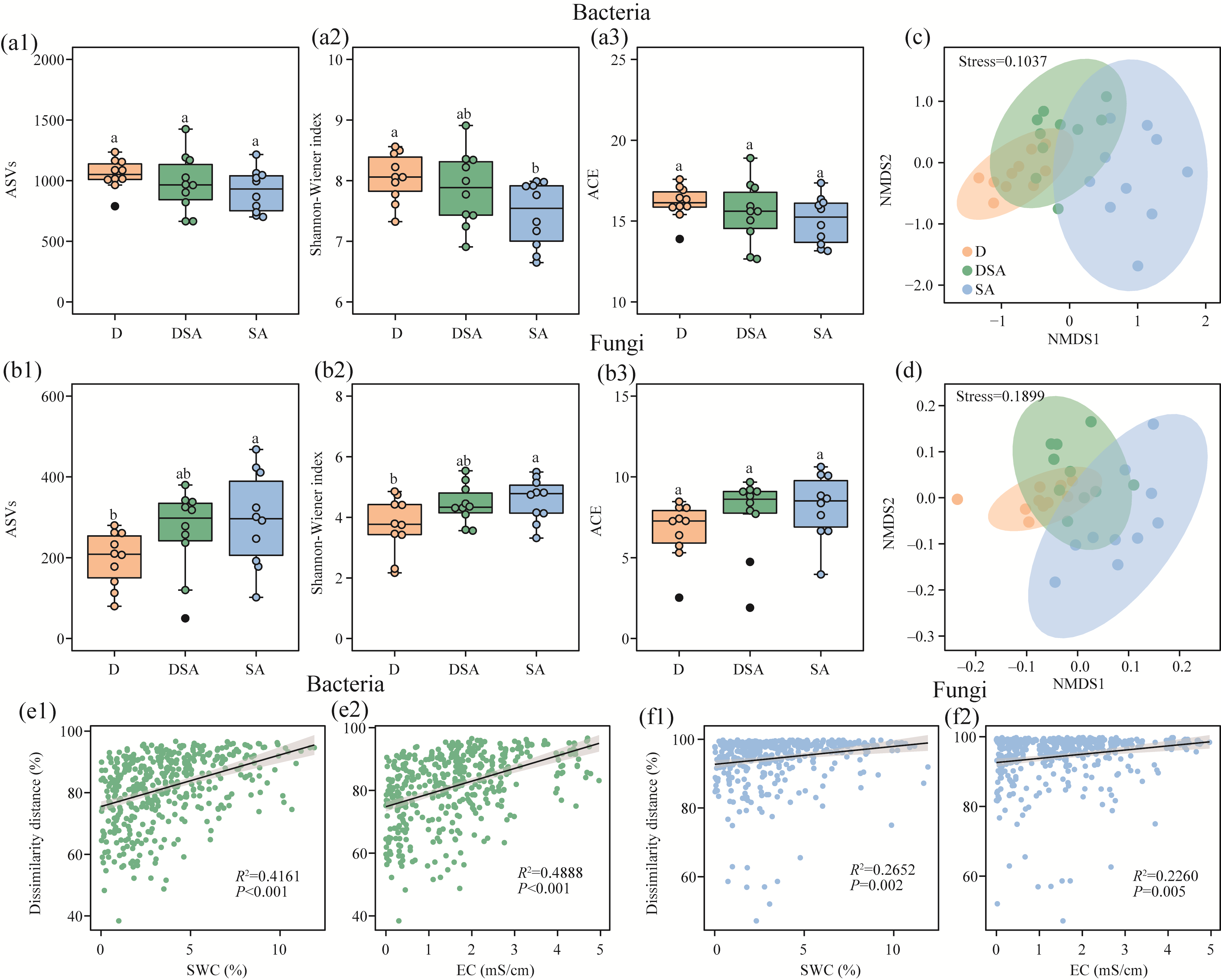
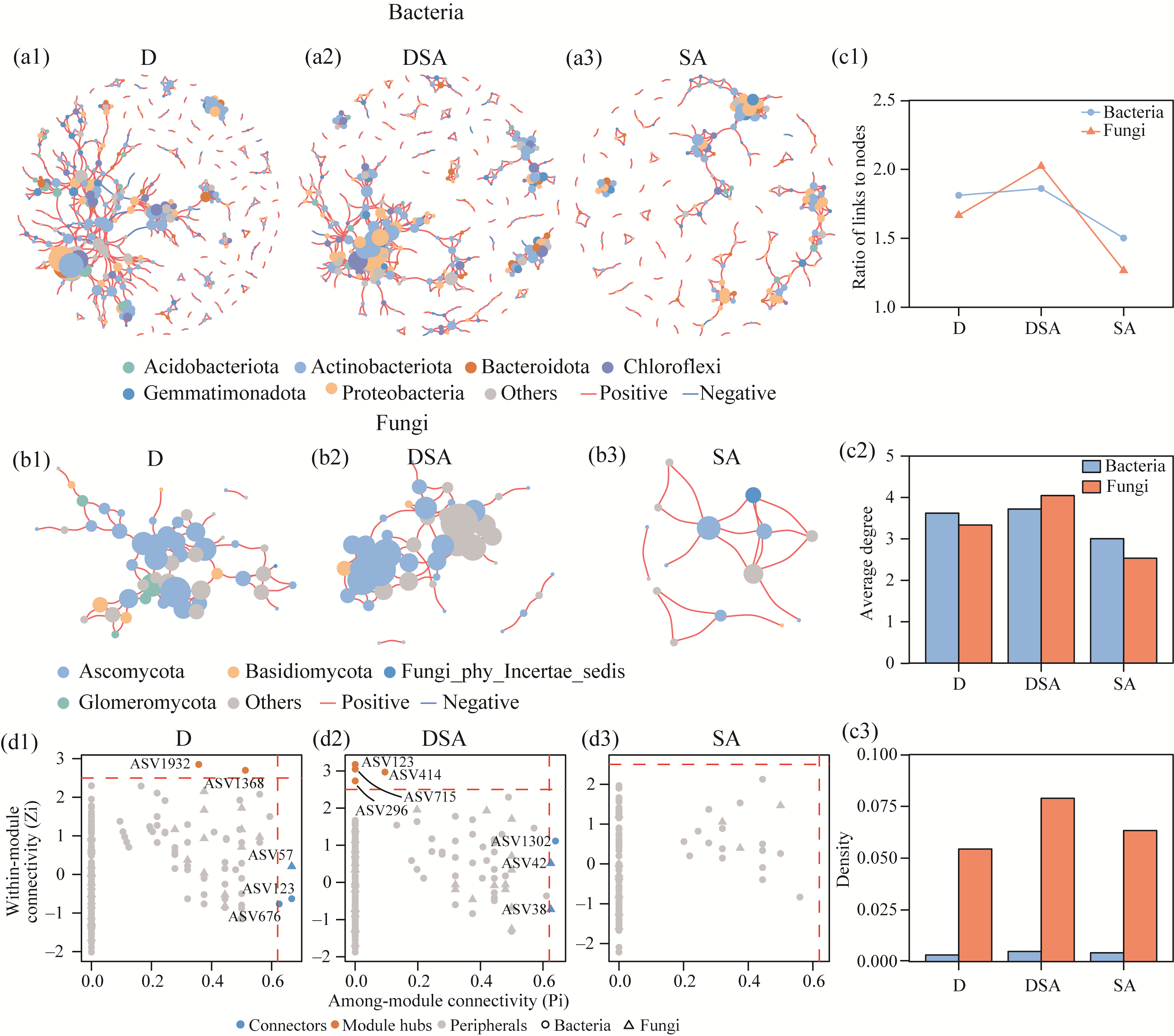
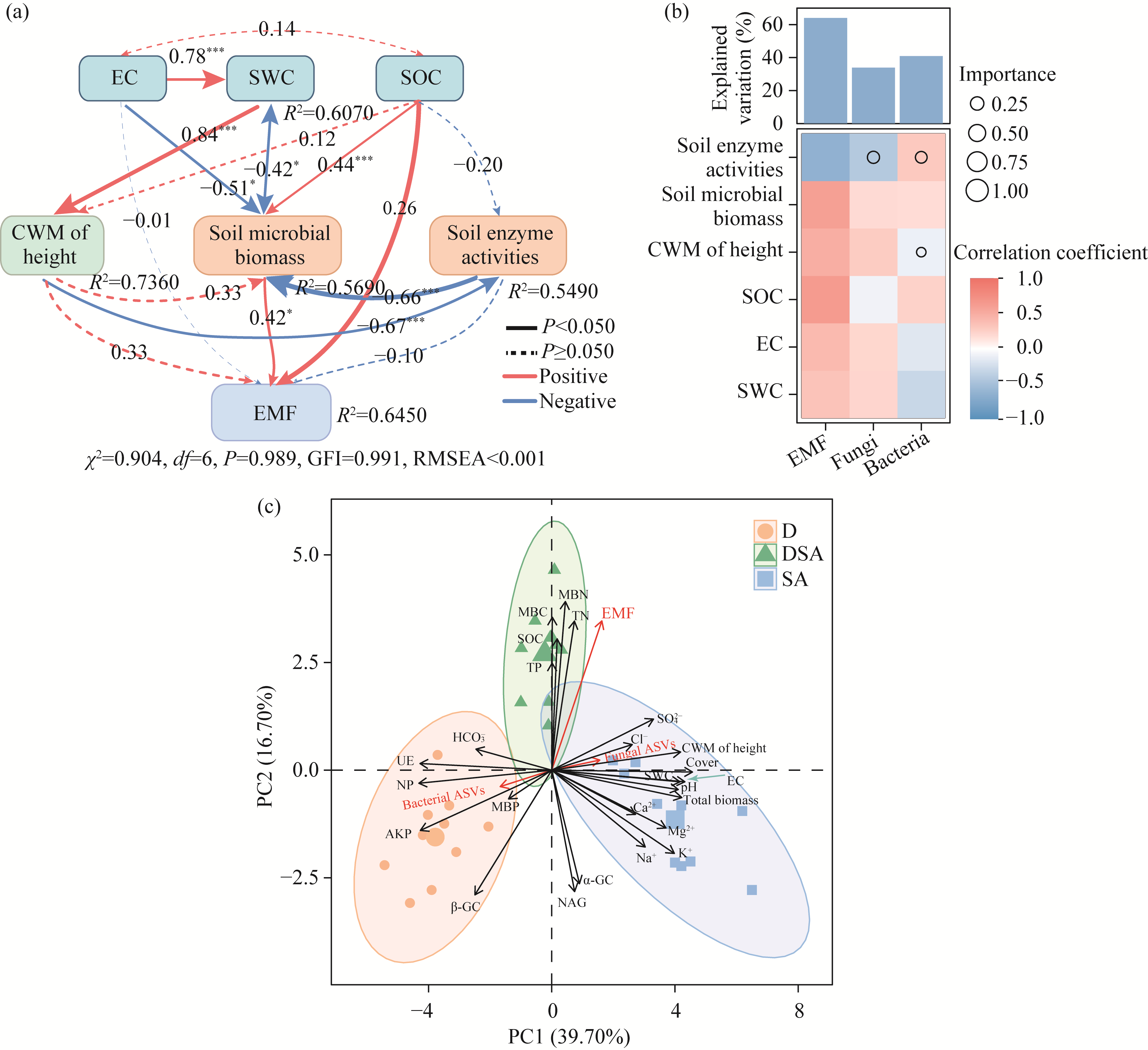
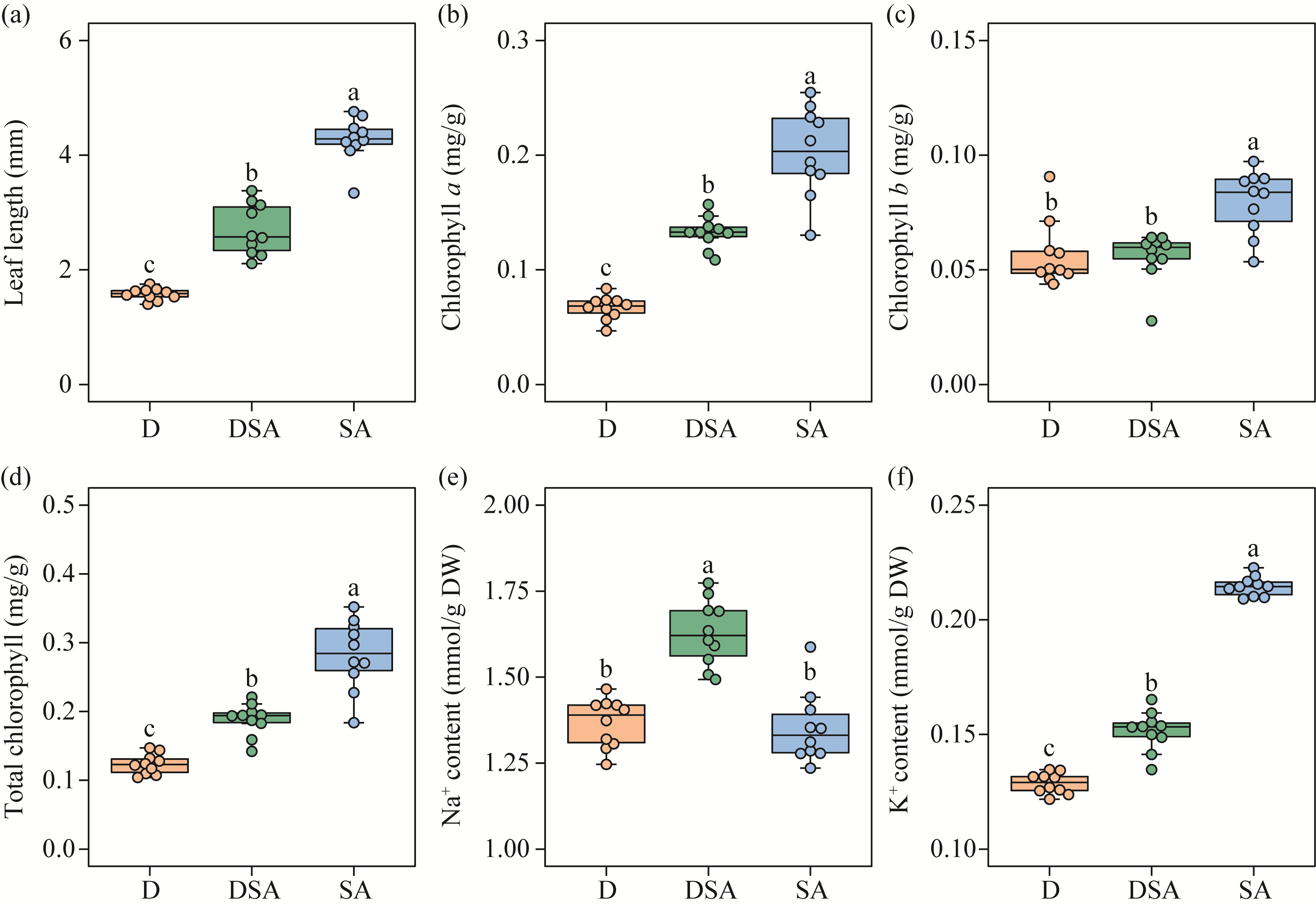
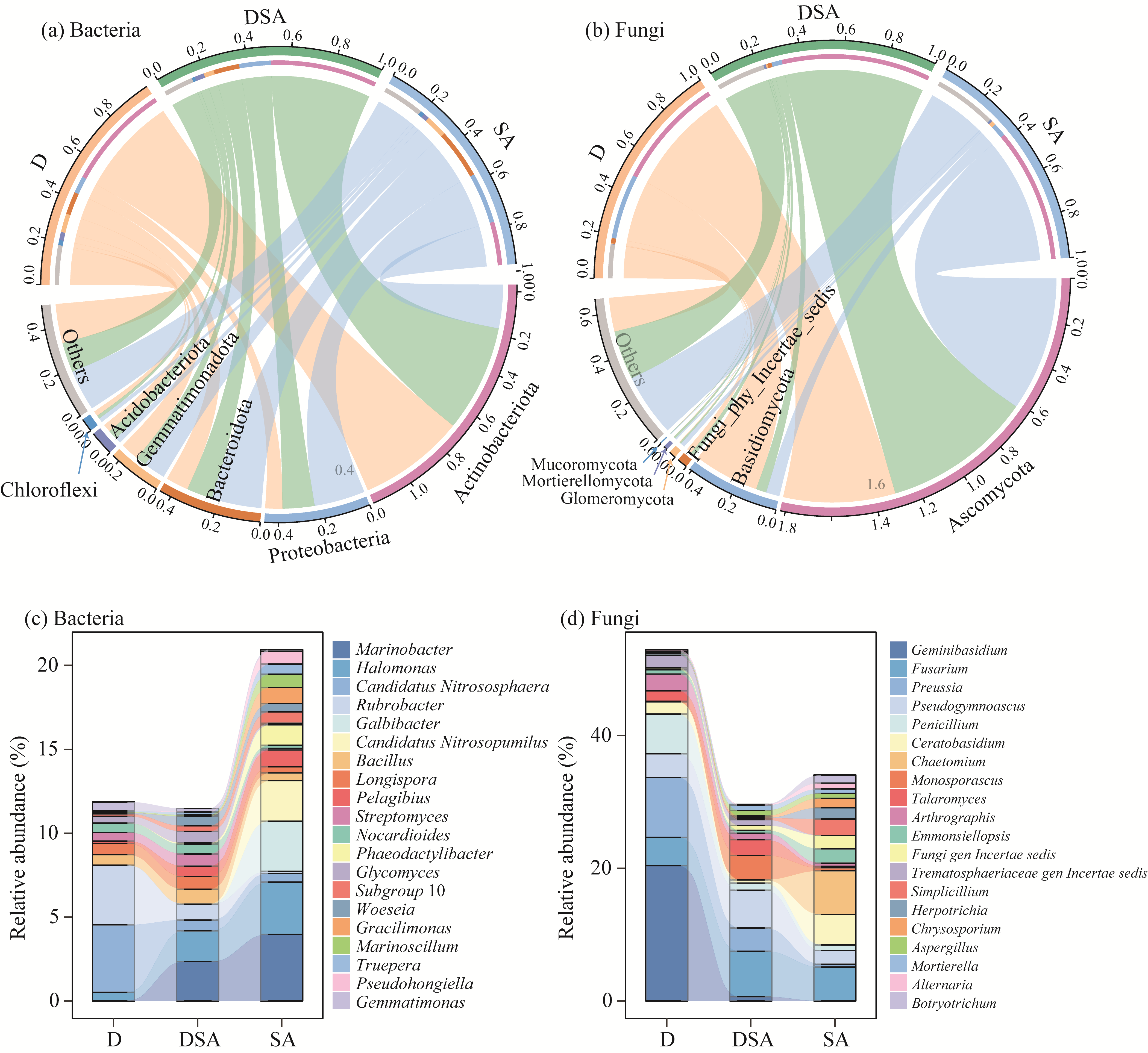
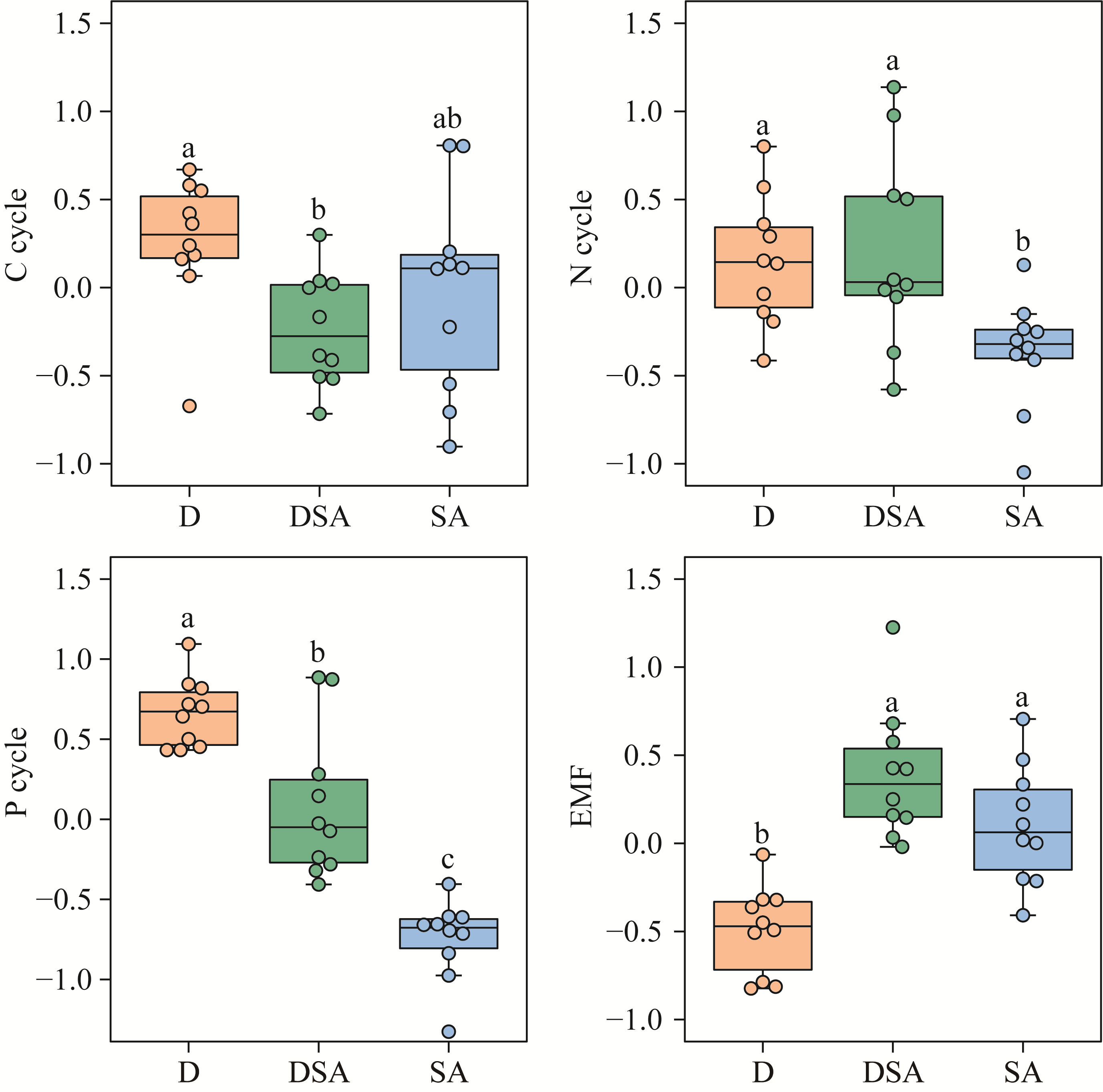
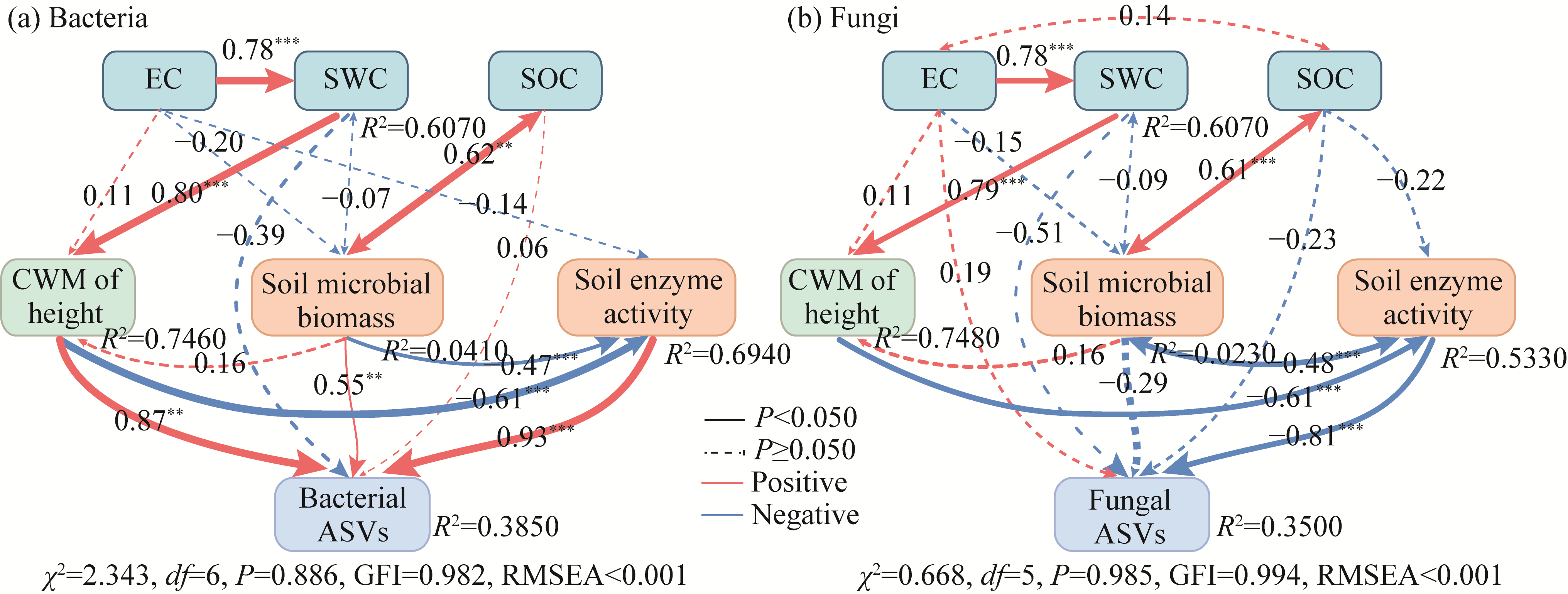
Abstract Soil water content and salinity critically regulate soil microbial composition, plant community structure, and ecosystem multifunctionality (EMF) in semi-arid grasslands. However, the mechanisms through which drought (D), saline-alkaline (SA), and their combined (DSA) stress influence these ecological components remain poorly understood. This study investigated these mechanisms along natural gradients in a semi-arid grassland of China by analyzing soil physical-chemical properties, microbial communities, and vegetation characteristics. The results showed that as the environmental stress shifted from the D group to the DSA group and then to the SA group, soil electrical conductivity significantly increased, while urease and phosphatase activities significantly decreased. Soil organic carbon, total nitrogen, total phosphorus, and microbial biomass carbon and nitrogen were lower in the D and SA groups than in the DSA group. Meanwhile, plant biomass showed an increasing trend along the treatment gradient, primarily driven by dominant species, while plant diversity did not exhibit significant differences. Further analysis identified the soil water content and salinity as the key determinants of soil microbial diversity and community complexity. Soil enzyme activities exhibited contrasting relationships with microbial composition, correlating positively with the richness of bacterial amplicon sequence variants (ASVs) but negatively with the richness of fungal ASVs. Notably, microbial biomass, which varied significantly across different groups, emerged as a key predictor of changes in EMF, with its critical role confirmed through structural equation modeling. These findings collectively elucidate the responses of ecological communities to synergistic soil hydro-saline stress in semi-arid ecosystems, while highlighting the critical role of microbial biomass in maintaining EMF.
|
|
Received: 11 August 2025
Published: 31 March 2026
|
|
Corresponding Authors:
*ZHANG Jinlin (E-mail: jlzhang@lzu.edu.cn)
|
|
|
| [1] |
Ahluwalia O, Singh P C, Bhatia R. 2021. A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resources, Environment and Sustainability, 5: 100032, doi: 10.1016/j.resenv.2021.100032.
|
|
|
| [2] |
Bahram M, Hildebrand F, Forslund S K, et al. 2018. Structure and function of the global topsoil microbiome. Nature, 560: 233-237.
doi: 10.1038/s41586-018-0386-6
|
|
|
| [3] |
Bais H P, Weir T L, Perry L G, et al. 2006. The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Review of Plant Biology, 57: 233-266.
pmid: 16669762
|
|
|
| [4] |
Banerjee S, Schlaeppi K,van der Heijden M G A. 2018. Keystone taxa as drivers of microbiome structure and functioning. Nature Reviews Microbiology, 16: 567-576.
doi: 10.1038/s41579-018-0024-1
pmid: 29789680
|
|
|
| [5] |
Banwart S A, Nikolaidis N P, Zhu Y G, et al. 2019. Soil functions: Connecting Earth's critical zone. Annual Review of Earth and Planetary Sciences, 47: 333-359.
doi: 10.1146/annurev-earth-063016-020544
|
|
|
| [6] |
Chai Y N, Schachtman D P. 2022. Root exudates impact plant performance under abiotic stress. Trends in Plant Science, 27: 80-91.
doi: 10.1016/j.tplants.2021.08.003
|
|
|
| [7] |
Cheeseman J M. 2015. The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytologist, 206(2): 557-570.
doi: 10.1111/nph.13217
pmid: 25495078
|
|
|
| [8] |
Chen C, Yin G Y, Hou L J, et al. 2023. Reclamation of tidal flats to paddy soils reshuffles the soil microbiomes along a 53-year reclamation chronosequence: Evidence from assembly processes, co-occurrence patterns and multifunctionality. Environment International, 179: 108151, doi: 10.1016/j.envint.2023.108151.
|
|
|
| [9] |
Chen H H, Ma K Y, Huang Y, et al. 2022. Significant response of microbial community to increased salinity across wetland ecosystems. Geoderma, 415: 115778, doi: 10.1016/j.geoderma.2022.115778.
|
|
|
| [10] |
Chen Q L, Ding J, Zhu Y G, et al. 2020. Soil bacterial taxonomic diversity is critical to maintaining the plant productivity. Environment International, 140: 105766, doi: 10.1016/j.envint.2020.105766.
|
|
|
| [11] |
Cheng Y X, Zhang C Y, Zhao X H, et al. 2018. Biomass-dominant species shape the productivity-diversity relationship in two temperate forests. Annals of Forest Science, 75(4): 97, doi: 10.1007/s13595-018-0780-0.
|
|
|
| [12] |
Coban O,De Deyn G B, van der Ploeg M. 2022. Soil microbiota as game-changers in restoration of degraded lands. Science, 375(6584): abe0725, doi: 10.1126/science.abe0725.
|
|
|
| [13] |
Cong H J. 2018. Analysis of soil moisture under the condition of different fertilization in gully area of the Loess Plateau. Bulletin of Science and Technology, 34(5): 70-73. (in Chinese)
|
|
|
| [14] |
de Vries F T, Griffiths R I, Bailey M, et al. 2018. Soil bacterial networks are less stable under drought than fungal networks. Nature Communications, 9: 3033, doi: 10.1038/s41467-018-05516-7.
pmid: 30072764
|
|
|
| [15] |
Delgado-Baquerizo M, Oliverio A M, Brewer T E, et al. 2018. A global atlas of the dominant bacteria found in soil. Science, 359(6373): 320-325.
doi: 10.1126/science.aap9516
pmid: 29348236
|
|
|
| [16] |
FAO (Food and Agriculture Organization of the United Nations). 2021. Global map of salt-affected soils: GSASmap v1.0. [2025-05-03]. https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/
|
|
|
| [17] |
Fu B J, Stafford-Smith M, Fu C. 2021. Editorial overview: Dryland social-ecological systems in changing environments. Current Opinion in Environmental Sustainability, 48: A1-A5.
|
|
|
| [18] |
Gao C, Xu L, Montoya L, et al. 2022. Co-occurrence networks reveal more complexity than community composition in resistance and resilience of microbial communities. Nature Communications, 13: 3867, doi: 10.1038/s41467-022-31343-y.
pmid: 35790741
|
|
|
| [19] |
Guan Y P, Jiang N N, Wu Y X, et al. 2021. Disentangling the role of salinity-sodicity in shaping soil microbiome along a natural saline-sodic gradient. Science of the Total Environment, 765: 142738, doi: 10.1016/j.scitotenv.2020.142738.
|
|
|
| [20] |
Hartmann M, Six J. 2023. Soil structure and microbiome functions in agroecosystems. Nature Reviews Earth & Environment, 4: 4-18.
|
|
|
| [21] |
Hooper D U, Adair E C, Cardinale B J, et al. 2012. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature, 486: 105-108.
doi: 10.1038/nature11118
|
|
|
| [22] |
Hu J P, Zhang M X, Lü Z L, et al. 2023. Grazing practices affect phyllosphere and rhizosphere bacterial communities of Kobresia humilis by altering their network stability. Science of The Total Environment, 900: 165814, doi: 10.1016/j.scitotenv.2023.165814.
|
|
|
| [23] |
Hu J P, He Y Y, Li J H, et al. 2024. Planting halophytes increases the rhizosphere ecosystem multifunctionality via reducing soil salinity. Environmental Research, 261: 119707, doi: 10.1016/j.envres.2024.119707.
|
|
|
| [24] |
Hu W G, Ran J Z, Dong L W, et al. 2021. Aridity-driven shift in biodiversity-soil multifunctionality relationships. Nature Communications, 12: 5350, doi: 10.1038/s41467-021-25641-0.
pmid: 34504089
|
|
|
| [25] |
Jansson J K, McClure R, Egbert R G. 2023. Soil microbiome engineering for sustainability in a changing environment. Nature Biotechnology, 41: 1716-1728.
doi: 10.1038/s41587-023-01932-3
pmid: 37903921
|
|
|
| [26] |
Ji L, Tian C J, Kuramae E E. 2023. Phosphorus-mediated succession of microbial nitrogen, carbon, and sulfur functions in rice-driven saline-alkali soil remediation. Soil Biology and Biochemistry, 184: 109125, doi: 10.1016/j.soilbio.2023.109125.
|
|
|
| [27] |
Jiao S, Chu H Y, Zhang B G, et al. 2022. Linking soil fungi to bacterial community assembly in arid ecosystems. iMeta, 1(1): e2, doi: 10.1002/imt2.2.
|
|
|
| [28] |
Jing X, Sanders N J, Shi Y, et al. 2015. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nature Communications, 6: 8159, doi: 10.1038/ncomms9159.
pmid: 26328906
|
|
|
| [29] |
Kang P, Pan Y Q, Yang P, et al. 2022. A comparison of microbial composition under three tree ecosystems using the stochastic process and network complexity approaches. Frontiers in Microbiology, 13: 1018077, doi: 10.3389/fmicb.2022.1018077.
|
|
|
| [30] |
Kouřilová X, Schwarzerová J, Pernicová I, et al. 2021. The first insight into polyhydroxyalkanoates accumulation in multi- extremophilic Rubrobacter xylanophilus and Rubrobacter spartanus. Microorganisms, 9(5): 909, doi: 10.3390/microorganisms9050909.
|
|
|
| [31] |
Li C J, Fu B J, Wang S, et al. 2021. Drivers and impacts of changes in China's drylands. Nature Reviews Earth & Environment, 2: 858-873.
|
|
|
| [32] |
Li J G, Pu L J, Han M F, et al. 2014. Soil salinization research in China: Advances and prospects. Journal of Geographical Sciences, 24: 943-960.
doi: 10.1007/s11442-014-1130-2
|
|
|
| [33] |
Li Y, Li W J, Jiang L M, et al. 2024. Salinity affects microbial function genes related to nutrient cycling in arid regions. Frontiers in Microbiology, 15: 1407760, doi: 10.3389/fmicb.2024.1407760.
|
|
|
| [34] |
Maestre F T, Quero J L, Gotelli N J, et al. 2012. Plant species richness and ecosystem multifunctionality in global drylands. Science, 335(6065): 214-218.
doi: 10.1126/science.1215442
pmid: 22246775
|
|
|
| [35] |
Manning P, van der Plas F, Soliveres S, et al. 2018. Redefining ecosystem multifunctionality. Nature Ecology & Evolution, 2: 427-436.
|
|
|
| [36] |
Mao D H, Wang Z M, Wu B F, et al. 2018. Land degradation and restoration in the arid and semiarid zones of China: Quantified evidence and implications from satellites. Land Degradation & Development, 2911 3841-3851.
|
|
|
| [37] |
Mo K C, Lettenmaier D P. 2014. Objective drought classification using multiple land surface models. Journal of Hydrometeorology, 15(3): 990-1010.
doi: 10.1175/JHM-D-13-071.1
|
|
|
| [38] |
Muhammad M, Waheed A, Wahab A, et al. 2024. Soil salinity and drought tolerance: An evaluation of plant growth, productivity, microbial diversity, and amelioration strategies. Plant Stress, 11: 100319, doi: 10.1016/j.stress.2023.100319.
|
|
|
| [39] |
Niu G X, Hasi M, Wang R Z, et al. 2021. Soil microbial community responses to long-term nitrogen addition at different soil depths in a typical steppe. Applied Soil Ecology, 167: 104054, doi: 10.1016/j.apsoil.2021.104054.
|
|
|
| [40] |
Pan Y Q, Kang P, Hu J P, et al. 2021. Bacterial community demonstrates stronger network connectivity than fungal community in desert-grassland salt marsh. Science of the Total Environment, 798: 149118, doi: 10.1016/j.scitotenv.2021.149118.
|
|
|
| [41] |
Pereira T A, Vieira S A, Oliveira R S, et al. 2022. Local drivers of heterogeneity in a tropical forest: Epiphytic tank bromeliads affect the availability of soil resources and conditions and indirectly affect the structure of seedling communities. Oecologia, 199: 205-215.
doi: 10.1007/s00442-022-05179-8
pmid: 35526202
|
|
|
| [42] |
Philippot L, Chenu C, Kappler A, et al. 2024. The interplay between microbial communities and soil properties. Nature Reviews Microbiology, 22: 226-239.
doi: 10.1038/s41579-023-00980-5
|
|
|
| [43] |
Sheng D C, Liu T, Wang H Y, et al. 2024. Advancing the dominance of winter annuals under changing rainfall patterns in a temperate desert of Central Asia. Global Ecology and Conservation, 54: e03064, doi: 10.1016/j.gecco.2024.e03064.
|
|
|
| [44] |
Shu W S, Huang L N. 2022. Microbial diversity in extreme environments. Nature Reviews Microbiology, 20: 219-235.
doi: 10.1038/s41579-021-00648-y
|
|
|
| [45] |
Singh J S, Gupta V K. 2018. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Science of the Total Environment, 634: 497-500.
doi: 10.1016/j.scitotenv.2018.03.373
|
|
|
| [46] |
Sritongon N, Sarin P, Theerakulpisut P, et al. 2022. The effect of salinity on soil chemical characteristics, enzyme activity and bacterial community composition in rice rhizospheres in Northeastern Thailand. Scientific Reports, 12: 20360, doi: 10.1038/s41598-022-24902-2.
pmid: 36437295
|
|
|
| [47] |
Tilman D, Isbell F, Cowles J M. 2014. Biodiversity and ecosystem functioning. Annual Review of Ecology, Evolution, and Systematics, 45: 471-493.
doi: 10.1146/ecolsys.2014.45.issue-1
|
|
|
| [48] |
Token S, Jiang L, Zhang L, et al. 2022. Effects of plant diversity on primary productivity and community stability along soil water and salinity gradients. Global Ecology and Conservation, 36: e02095, doi: 10.1016/j.gecco.2022.e02095.
|
|
|
| [49] |
Trivedi P, Leach J E, Tringe S G, et al. 2020. Plant-microbiome interactions: from community assembly to plant health. Nature Reviews Microbiology, 18: 607-621.
doi: 10.1038/s41579-020-0412-1
|
|
|
| [50] |
Wagg C, Roscher C, Weigelt A, et al. 2022. Biodiversity-stability relationships strengthen over time in a long-term grassland experiment. Nature Communications, 13: 7752, doi: 10.1038/s41467-022-35189-2.
pmid: 36517483
|
|
|
| [51] |
Wan N F, Zheng X R, Fu L W, et al. 2020. Global synthesis of effects of plant species diversity on trophic groups and interactions. Nature Plants, 6: 503-510.
doi: 10.1038/s41477-020-0654-y
|
|
|
| [52] |
Wang C S, Wang H Q, Wang W, et al. 2022a. The salt secretion of leaves promotes the competitiveness of Reaumuria soongarica in a desert grassland. BMC Plant Biology, 22: 85, doi: 10.1186/s12870-022-03457-4.
|
|
|
| [53] |
Wang S, Wang X B, Han X G, et al. 2018. Higher precipitation strengthens the microbial interactions in semi-arid grassland soils. Global Ecology and Biogeography, 27(5): 570-580.
doi: 10.1111/geb.2018.27.issue-5
|
|
|
| [54] |
Wang W Y, Jia T H, Qi T Y, et al. 2022b. Root exudates enhanced rhizobacteria complexity and microbial carbon metabolism of toxic plants. iScience, 25(10): 105243, doi: 10.1016/j.isci.2022.105243.
|
|
|
| [55] |
Wang X, Zhang Q, Zhang Z J, et al. 2023. Decreased soil multifunctionality is associated with altered microbial network properties under precipitation reduction in a semiarid grassland. iMeta, 2(2): e106, doi: 10.1002/imt2.106.
|
|
|
| [56] |
Wei H H, Geng X Y, Zhu W, et al. 2023. Individual and combined influences of salinity and drought stress on the agro-physiological traits and grain yield of rice. Field Crops Research, 304: 109172, doi: 10.1016/j.fcr.2023.109172.
|
|
|
| [57] |
Wei Y X, Chen L J, Feng Q, et al. 2024. Structure and assembly mechanism of soil bacterial community under different soil salt intensities in arid and semiarid regions. Ecological Indicators, 158: 111631, doi: 10.1016/j.ecolind.2024.111631.
|
|
|
| [58] |
Wu L W, Zhang Y, Guo X, et al. 2022. Reduction of microbial diversity in grassland soil is driven by long-term climate warming. Nature Microbiology, 7: 1054-1062.
doi: 10.1038/s41564-022-01147-3
pmid: 35697795
|
|
|
| [59] |
Xi H Y, Feng Q, Zhang L, et al. 2016. Effects of water and salinity on plant species composition and community succession in Ejina Desert Oasis, northwest China. Environmental Earth Sciences, 75: 138, doi: 10.1007/s12665-015-4823-7.
|
|
|
| [60] |
Xiao Y, Liu X, Zhang L, et al. 2021. The allometry of plant height explains species loss under nitrogen addition. Ecology Letters, 24(3): 553-562.
doi: 10.1111/ele.13673
pmid: 33423373
|
|
|
| [61] |
Yan N, Marschner P, Cao W H, et al. 2015. Influence of salinity and water content on soil microorganisms. International Soil and Water Conservation Research, 3(4): 316-323.
doi: 10.1016/j.iswcr.2015.11.003
|
|
|
| [62] |
Yang G W, Roy J, Veresoglou S D, et al. 2021. Soil biodiversity enhances the persistence of legumes under climate change. New Phytologist, 229(5): 2945-2956.
doi: 10.1111/nph.17065
pmid: 33152109
|
|
|
| [63] |
Yao S R, Hu W G, Ji M F, et al. 2025. Distribution, species richness, and relative importance of different plant life forms across drylands in China. Plant Diversity, 47(2): 273-281.
doi: 10.1016/j.pld.2024.09.007
|
|
|
| [64] |
Ye H, Hong M, Xu X H, et al. 2024. Responses of plant diversity and soil microorganism diversity to nitrogen addition in the desert steppe, China. Journal of Arid Land, 16(3): 447-459.
doi: 10.1007/s40333-024-0008-0
|
|
|
| [65] |
Zhang G L, Bai J H, Zhai Y J, et al. 2024a. Microbial diversity and functions in saline soils: A review from a biogeochemical perspective. Journal of Advanced Research, 59: 129-140.
doi: 10.1016/j.jare.2023.06.015
|
|
|
| [66] |
Zhang J, Guo X Q, Shan Y J, et al. 2024b. Effects of land-use patterns on soil microbial diversity and composition in the Loess Plateau, China. Journal of Arid Land, 16(3): 415-430.
doi: 10.1007/s40333-024-0007-1
|
|
|
| [67] |
Zhang T J, Chen Y N, Ali S. 2023. Abiotic stress and human activities reduce plant diversity in desert riparian forests. Ecological Indicators, 152: 110340, doi: 10.1016/j.ecolind.2023.110340.
|
|
|
| [68] |
Zheng J H, Zhang B, Zhang F, et al. 2023. Effects of fencing on near-term ecosystem multifunctionality in a typical steppe in Inner Mongolia. Agriculture, Ecosystems & Environment, 342: 108238, doi: 10.1016/j.agee.2022.108238.
|
|
|
| [69] |
Zhou J Q, Gong J C, Wang P S, et al. 2024. Historical tillage promotes grass-legume mixtures establishment and accelerates soil microbial activity and organic carbon decomposition. Journal of Arid Land, 16(7): 910-924.
doi: 10.1007/s40333-024-0021-3
|
|
|
| [70] |
Zhou J Z, Ning D L. 2017. Stochastic community assembly: Does it matter in microbial ecology. Microbiology and Molecular Biology Reviews, 81(4): e00002-17, doi: 10.1128/MMBR.00002-17.
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|