| Research article |
|


|
|
|
| Plasticity of photorespiratory carbon concentration mechanism in Sedobassia sedoides (Pall.) Freitag & G. Kadereit under elevated CO2 concentration and salinity |
Zulfira RAKHMANKULOVA1, Elena SHUYSKAYA1, Maria PROKOFIEVA1, Kristina TODERICH2,*( )( )( ), Pavel VORONIN1 ), Pavel VORONIN1 |
1Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Moscow 127276, Russia
2Graduate School of Bioresources, Mie University, 1577 Kurimamachiya, Tsu 514-8507, Japan |
|
|
|
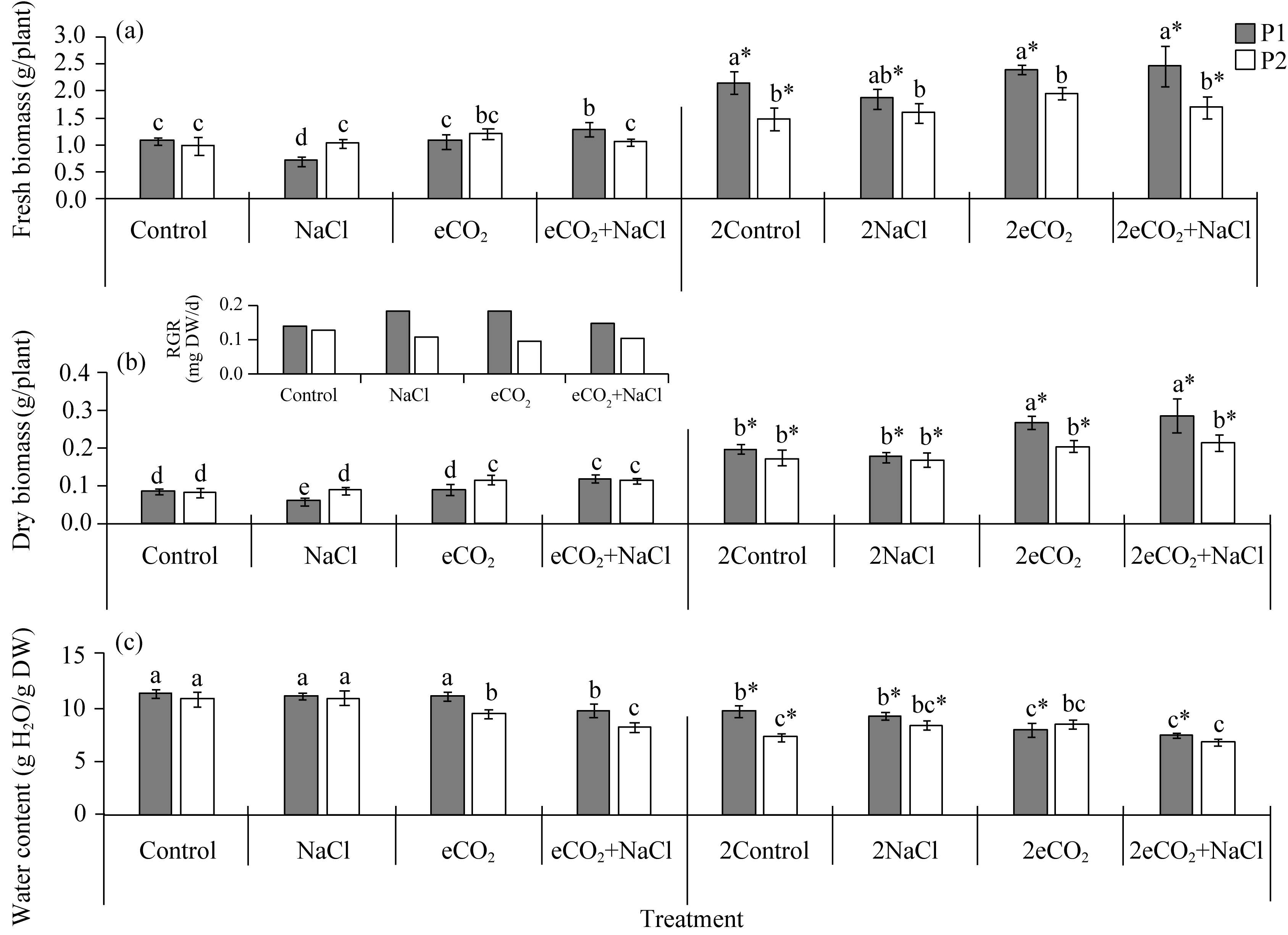
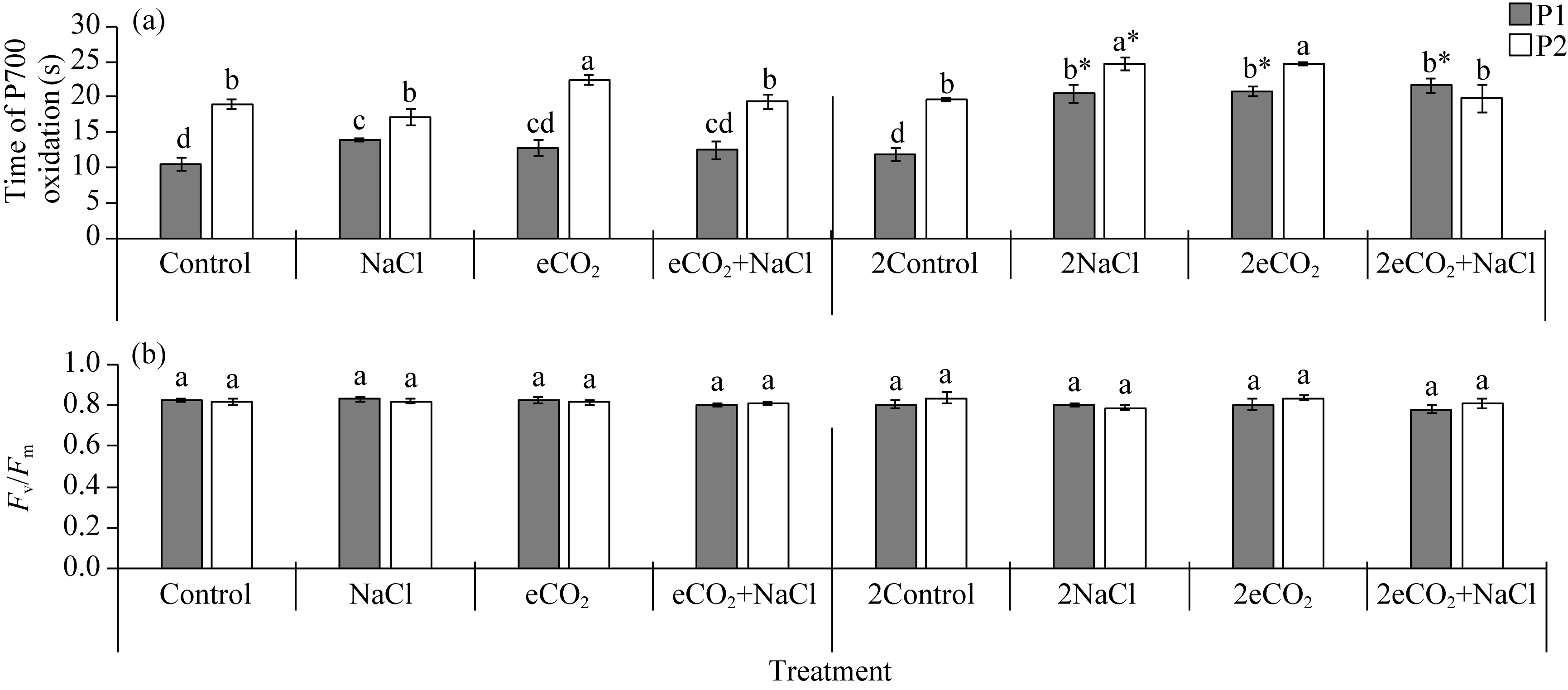
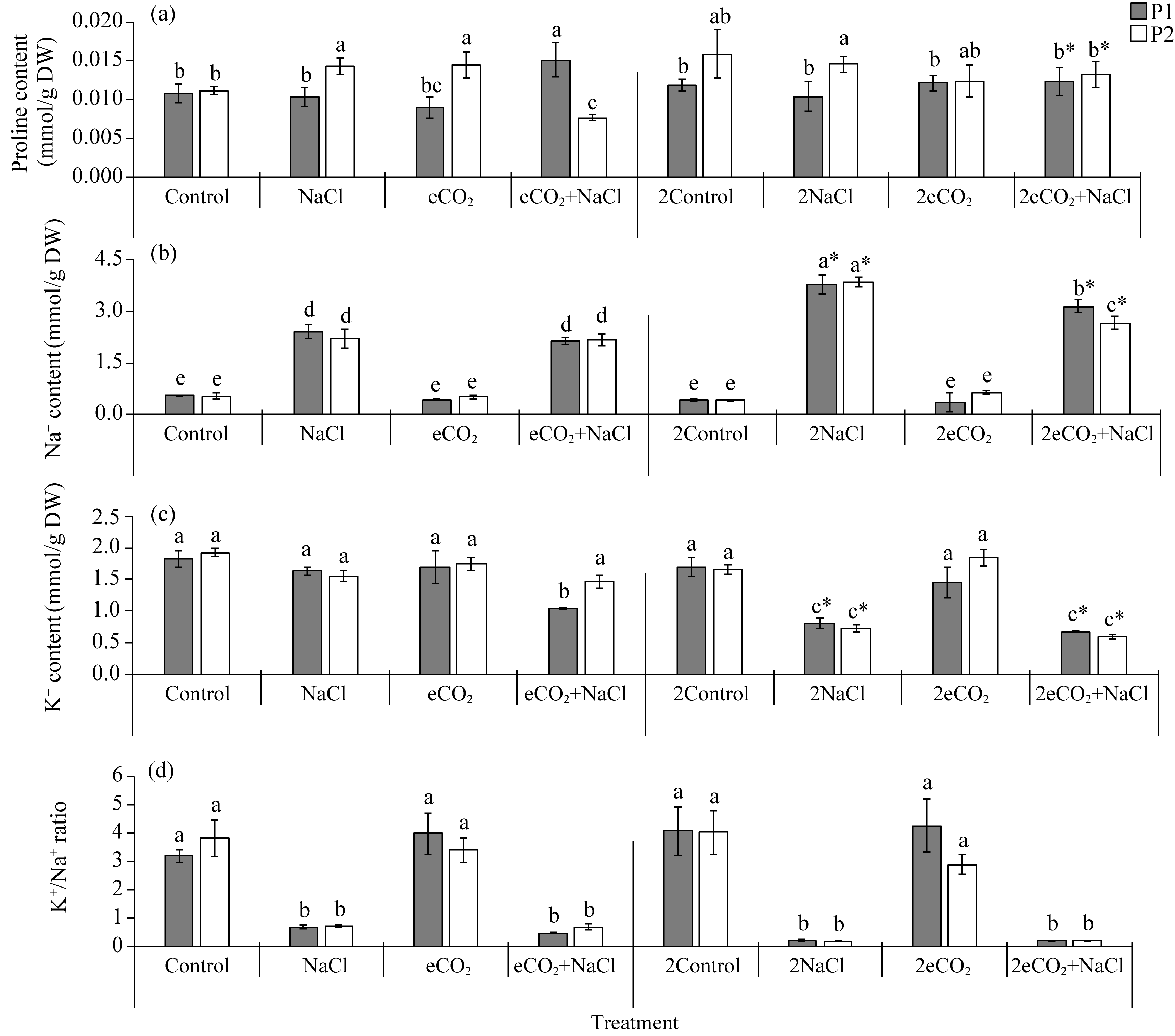
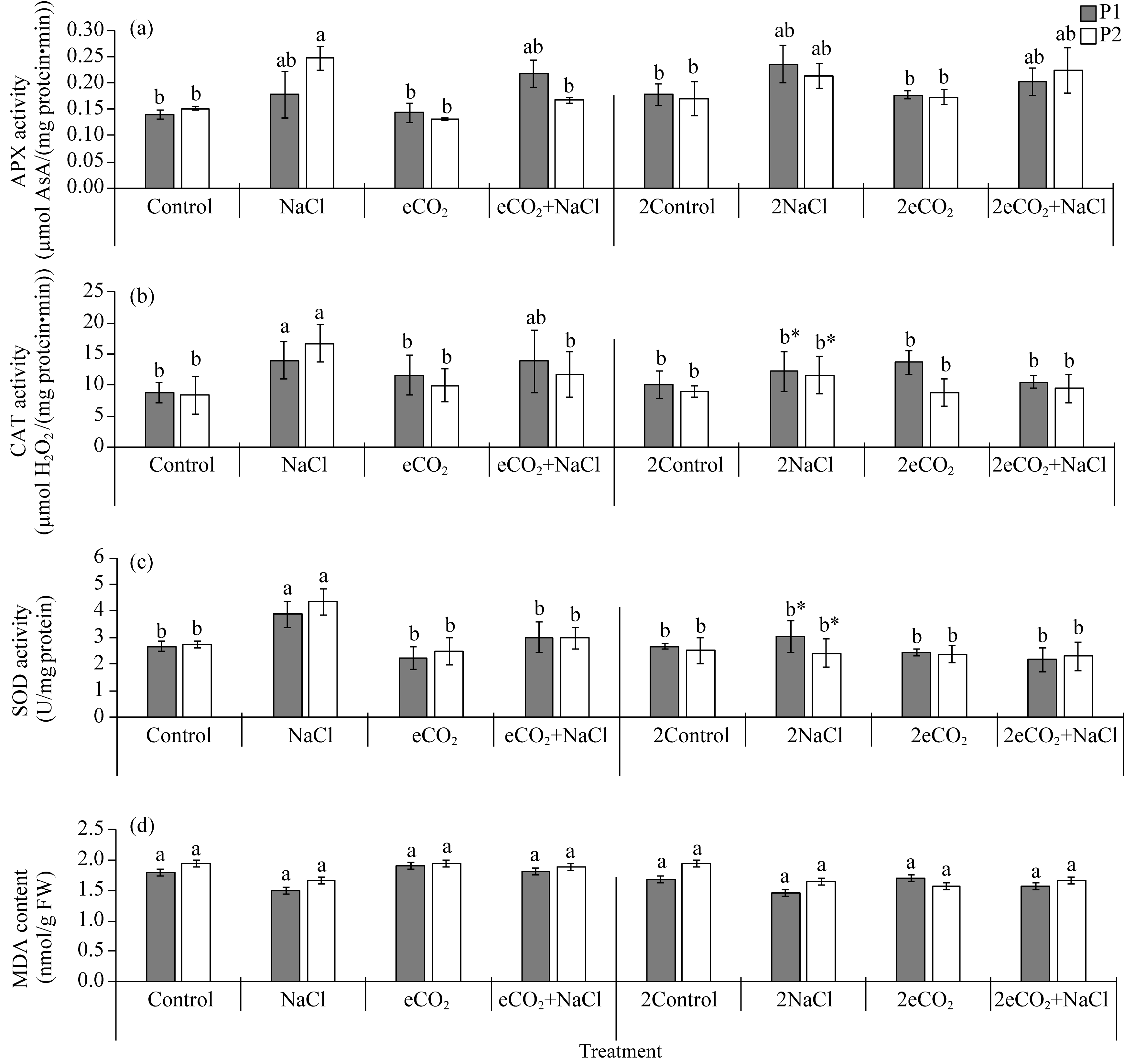
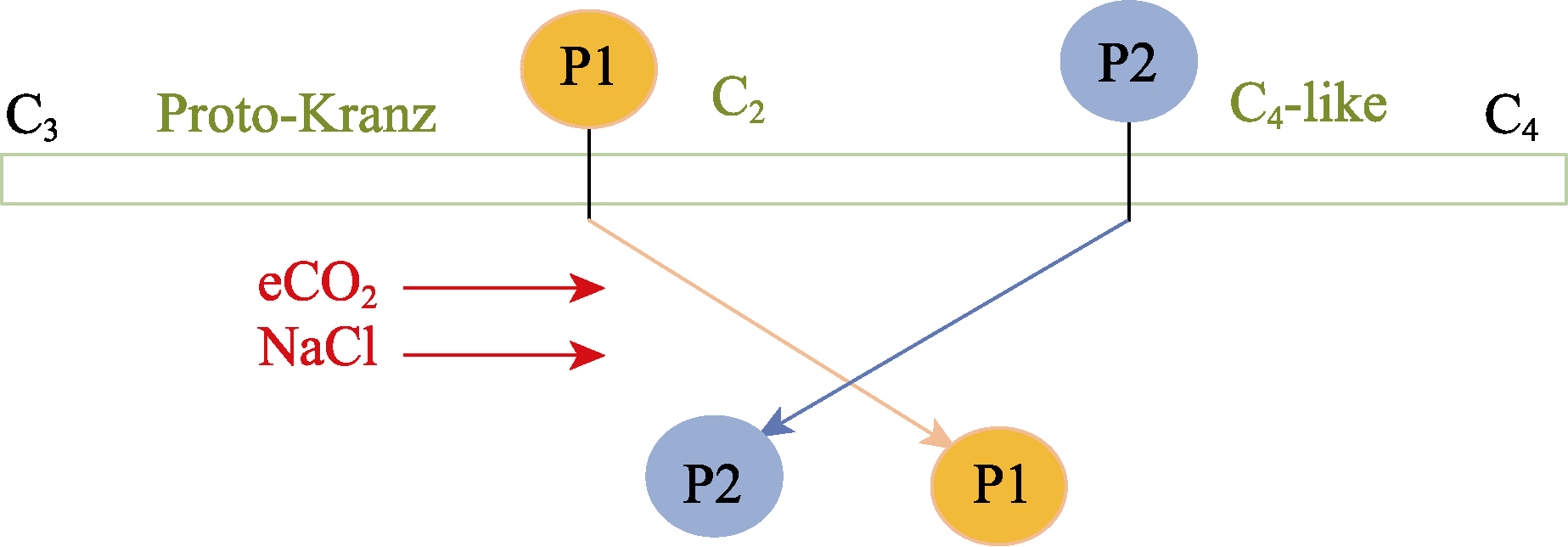
Abstract Rising atmospheric CO2 (carbon dioxide) concentrations and salinization are manifestations of climate change that affect plant growth and productivity. Species with an intermediate C3-C4 type of photosynthesis live in a wide range of precipitation, temperature, and soil quality, but are more often found in warm and dry habitats. One of the intermediate C3-C4 photosynthetic type is C2 photosynthesis with a carbon concentration mechanism (CCM) that reassimilates CO2 released via photorespiration. However, the ecological significance under which C2 photosynthesis has advantages over C3 and C4 plants remains largely unexplored. Salt tolerance and functioning of CCM were studied in plants from two populations (P1 and P2) of Sedobassia sedoides (Pall.) Freitag & G. Kadereit Asch. species with C2 photosynthesis exposed to 4 d and 10 d salinity (200 mM NaCl) at ambient (785.7 mg/m3, aCO2) and elevated (1571.4 mg/m3, eCO2) CO2. On the fourth day of salinity, an increase in Na+ content, activity catalase, and superoxide dismutase was observed in both populations. P2 plants showed an increase in proline content and a decrease in photosynthetic enzyme content: rubisco, phosphoenolpyruvate carboxylase (PEPC), and glycine decarboxylase (GDC), which indicated a weakening of C2 and C4 characteristics under salinity. Treatment under 10 d salinity led to an increased Na+ content and activity of cyclic electron flow around photosystem I (PSI CEF), a decreased content of K+ and GDC in both populations. P1 plants showed greater salt tolerance, which was assessed by the degree of reduction in photosynthetic enzyme content, PSI CEF activity, and changes in relative growth rate (RGR). Differences between populations were evident under the combination of eCO2 and salinity. Under long-term salinity and eCO2, more salt-tolerant P1 plants had a higher dry biomass (DW), which was positively correlated with PSI CEF activity. In less salt-tolerant P2 plants, DW correlated with transpiration and dark respiration. Thus, S. sedoides showed a high degree of photosynthetic plasticity under the influence of salinity and eCO2 through strengthening (P1 plants) and weakening C4 characteristics (P2 plants).
|
|
Received: 09 February 2024
Published: 31 July 2024
|
|
Corresponding Authors:
* Kristina TODERICH (E-mail: ktoderich@tottori-u.ac.jp, ktoderich@bio.mie-u.ac.jp; ORCID: 0000-0003-2825-7214)
|
|
|
| [1] |
Aebi H. 1984. Catalase in vitro. Methods Enzymology, 105: 121-126.
|
|
|
| [2] |
Allakhverdiev S I, Nishiyama Y, Miyairi S, et al. 2002. Salt stress inhibits the repair of photodamaged photosystem II by suppressing the transcription and translation of psbA genes in Synechocystis. Plant Physiology, 130(3): 1433-1443.
|
|
|
| [3] |
Aranda-Sicilia M N, Aboukila A, Armbruster U, et al. 2016. Envelope K+/H+ antiporters AtKEA1 and AtKEA 2 function in plastid development. Plant Physiology, 172(1): 441-449.
doi: 10.1104/pp.16.00995
pmid: 27443603
|
|
|
| [4] |
Bai J H, Qin Y, Liu J H, et al. 2017. Proteomic response of oat leaves to long-term salinity stress. Environmental Science and Pollution Research, 24(4): 3387-3399.
|
|
|
| [5] |
Bates L S, Waldren R P, Teare I D. 1973. Rapid determination of free proline for water stress studies. Plant and Soil, 39: 205-207.
|
|
|
| [6] |
Beauchamp C, Fridovich I. 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analytical Biochemistry, 44(1): 276-287.
doi: 10.1016/0003-2697(71)90370-8
pmid: 4943714
|
|
|
| [7] |
Bose J, Rodrigo-Moreno A, Shabala S. 2014. ROS homeostasis in halophytes in the context of salinity stress tolerance. Journal of Experimental Botany, 65(5): 1241-1257.
doi: 10.1093/jxb/ert430
pmid: 24368505
|
|
|
| [8] |
Bose J, Munns R, Shabala S, et al. 2017. Chloroplast function and ion regulation in plants growing on saline soils: Lessons from halophytes. Journal of Experimental Botany, 68(12): 3129-3143.
doi: 10.1093/jxb/erx142
pmid: 28472512
|
|
|
| [9] |
Bradford M M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72(1-2): 248-254.
|
|
|
| [10] |
Chaves M, Flexas J, Pinheiro C. 2009. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Annals of Botany, 103(4): 551-560.
doi: 10.1093/aob/mcn125
pmid: 18662937
|
|
|
| [11] |
Chen J, Wang P, Mi H L, et al. 2010. Reversible association of ribulose-1, 5-bisphosphate carboxylase/oxygenase activase with the thylakoid membrane depends upon the ATP level and pH in rice without heat stress. Journal of Experimental Botany, 61(11): 2939-2950.
doi: 10.1093/jxb/erq122
pmid: 20478969
|
|
|
| [12] |
Chen T, Wang W, Xu K, et al. 2019. K+ and Na+ transport contribute to K+/Na+ homeostasis in Pyropia haitanensis under hypersaline stress. Algal Research, 40: 101526, doi: 10.1016/j.algal.2019.101526.
|
|
|
| [13] |
Ellouzi H, Hamed K B, Cela J, et al. 2011. Early effects of salt stress on the physiological and oxidative status of Cakile maritima (halophyte) and Arabidopsis thaliana (glycophyte). Physiologia Plantarum, 142(2): 128-143.
|
|
|
| [14] |
Fricke W, Akhiyarova G, Veselov D, et al. 2004. Rapid and tissue-specific changes in ABA and in growth rate in response to salinity in barley leaves. Journal of Experimental Botany, 55(399): 1115-1123.
pmid: 15047763
|
|
|
| [15] |
Goncharenko G G, Padutov V E, Silin A E. 1994. Construction of genetic maps for some Eurasian coniferous species using allozyme genes. Biochemical Genetics, 32: 223-236.
pmid: 7993376
|
|
|
| [16] |
Heath R L, Pasker L. 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Archives of Biochemistry and Biophysics, 125(1): 189-198.
doi: 10.1016/0003-9861(68)90654-1
pmid: 5655425
|
|
|
| [17] |
Johnson J E, Field C B, Berry J A. 2021. The limiting factors and regulatory processes that control the environmental responses of C3, C3-C4 intermediate, and C4 photosynthesis. Oecologia, 197: 841-866.
|
|
|
| [18] |
Jothiramshekar S, Benjamin J J, Krishnasamy R, et al. 2018. Responses of selected C3 and C4 halophytes to elevated CO2 concentration under salinity. Current Science, 115(1): 129-135.
|
|
|
| [19] |
Klughammer C, Schreiber U. 1998. Measuring P700 absorbance changes in the near infrared spectral region with a dual wavelength pulse modulation system. In: GarabG. Photosynthesis:Mechanisms and Effects. Dordrecht: Kluwer Academic Publishers, 4357-4360.
|
|
|
| [20] |
Konyushkova M V, Kozlov D N. 2010. Automated analysis of the distribution of dark-colored chernozemlike soils in the Northern Caspian region based on satellite imaging data: The example of the Dzhanybek station, Arid Ecosystems, 16(5): 46-56. (in Japanese)
|
|
|
| [21] |
Laemmli U K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
|
|
|
| [22] |
Lauterbach M, Schmidt H, Billakurthi K, et al. 2017. De novo transcriptome assembly and comparison of C3, C3-C4, and C4 species of tribe Salsoleae (Chenopodiaceae). Frontiers of Plant Science, 8: 1939, doi: 10.3389/fpls.2017.01939.
|
|
|
| [23] |
Lundgren M R, Christin P A, Escobar E G, et al. 2016. Evolutionary implications of C3-C4 intermediates in the grass Alloteropsis semialata. Plant, Cell and Environment, 39(9): 1874-1885.
|
|
|
| [24] |
Lundgren M R, Christin P A. 2017. Despite phylogenetic effects, C3-C4 lineages bridge the ecological gap to C4 photosynthesis. Journal of Experimental Botany, 68(2): 241-254.
|
|
|
| [25] |
Lundgren M R. 2020. C2 photosynthesis: A promising route towards crop improvement? New Phytologist, 228(6): 1734-1740.
|
|
|
| [26] |
Malinovsky A V, Akanov E N, Voronin P Y. 2020. A vegetation climatic unit for studying the impact on higher plants of an increased CO2 concentration in comparison with the atmospheric CO2 concentration. Russian Journal of Plant Physiology, 67: 194-200.
|
|
|
| [27] |
Mallmann J, Heckmann D, Bräutigam A, et al. 2014. The role of photorespiration during the evolution of C4 photosynthesis in the genus Flaveria. eLife, 3: e02478, doi: 10.7554/eLife.02478.
|
|
|
| [28] |
Munekage Y N, Taniguchi Y Y. 2022. A scheme for C4 evolution derived from a comparative analysis of the closely related C3, C3-C4 intermediate, C4-like, and C4 species in the genus Flaveria. Plant Molecular Biology, 110: 445-454.
doi: 10.1007/s11103-022-01246-z
pmid: 35119574
|
|
|
| [29] |
Nakamura N, Iwano M, Havaux M, et al. 2013. Promotion of cyclic electron transport around photosystem I during the evolution of NADP-malic enzyme-type C4 photosynthesis in the genus Flaveria. New Phytologist, 199(3): 832-842.
doi: 10.1111/nph.12296
pmid: 23627567
|
|
|
| [30] |
Nakano Y, Asada K. 1981. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant and Cell Physiology, 22(5): 867-880.
|
|
|
| [31] |
Nei M. 1987. Molecular Evolutionary Genetics. New York: Columbia University Press, 512.
|
|
|
| [32] |
Pan T, Liu M, Kreslavski V D, et al. 2020. Non-stomatal limitation of photosynthesis by soil salinity, Critical Reviews in Environmental Science and Technology, 51(8): 791-825.
|
|
|
| [33] |
Panta S, Flowers T, Lane P, et al. 2014. Halophyte agriculture: Success stories. Environmental and Experimental Botany, 107: 71-83.
|
|
|
| [34] |
Percey W J, McMinn A, Bose J, et al. 2016a. Salinity effects on chloroplast PSII performance in glycophytes and halophytes. Functional Plant Biology, 43(11): 1003-1015.
|
|
|
| [35] |
Percey W J, Shabala L, Wu Q, et al. 2016b. Potassium retention in leaf mesophyll as an element of salinity tissue tolerance in halophytes. Plant Physiology and Biochemistry, 109: 346-354.
|
|
|
| [36] |
Pérez-López U, Robredo A, Lacuesta M, et al. 2012. Elevated CO2 reduces stomatal and metabolic limitations on photosynthesis caused by salinity in Hordeum vulgare. Photosynthesis Research, 111(3): 269-283.
doi: 10.1007/s11120-012-9721-1
pmid: 22286185
|
|
|
| [37] |
Pérez-Romero J A, Idaszkin Y L, Barcia-Piedras J M, et al. 2018. Disentangling the effect of atmospheric CO2 enrichment on the halophyte Salicornia ramosissima J. woods physiological performance under optimal and suboptimal saline conditions. Plant Physiology and Biochemistry, 127: 617-629.
doi: S0981-9428(18)30199-2
pmid: 29738990
|
|
|
| [38] |
Rakhmankulova Z F, Shuyskaya E V, Suyundukov Y T, et al. 2016. Different responses of two ecotypes of C3-C4 xero-halophyte Bassia sedoides to osmotic and ionic factors of salt stress. Russian Journal of Plant Physiology, 63: 349-357.
|
|
|
| [39] |
Rakhmankulova Z F. 2018. Photorespiration: Its role in the productive process and evolution of С4 plants. Russian Journal of Plant Physiology, 65: 303-318.
|
|
|
| [40] |
Rakhmankulova Z F. 2022. Plant respiration and global climatic changes. Russian Journal of Plant Physiology, 69: 572-588.
|
|
|
| [41] |
Rakhmankulova Z F, Shuyskaya E V, Prokofieva M Y. 2023. Intraspecific photosynthetic diversity and differences in stress-induced plasticity in С3-С4 Sedobassia sedoides under drought stress. Russian Journal of Plant Physiology, 70: 81, doi: 10.1134/S1021443722603135.
|
|
|
| [42] |
Sage R F, Sage T L, Kocacinar F. 2012. Photorespiration and the evolution of C4 photosynthesis. Annual Review of Plant Biology, 63: 19-47.
|
|
|
| [43] |
Sage R F, Khoshravesh R, Sage T L. 2014. From proto-Kranz to C4 Kranz: Building the bridge to C4 photosynthesis. Journal of Experimental Botany, 65(13): 3341-3356.
|
|
|
| [44] |
Salmon Y, Lintunen A, Dayet A, et al. 2020. Leaf carbon and water status control stomatal and nonstomatal limitations of photosynthesis in trees. New Phytologist, 226(3): 690-703.
doi: 10.1111/nph.16436
pmid: 31955422
|
|
|
| [45] |
Sapanov M K, Sizemskaya M L. 2015. Climate change and dynamics of virgin land vegetation in the northern Caspian region. Povolzhskiy Journal of Ecology, 3: 307-320.
|
|
|
| [46] |
Schlüter U, Bouvier J W, Guerreiro R, et al. 2023. Brassicaceae display variation in efficiency of photorespiratory carbon- recapturing mechanisms. Journal of Experimental Botany, 74(21): 6631-6649.
doi: 10.1093/jxb/erad250
pmid: 37392176
|
|
|
| [47] |
Schreiber U, Gademann R, Ralph P J, et al. 1997. Assessment of photosynthetic performance of Prochloron in Lissoclinum patella in hospite by chlorophyll fluorescence measurements. Plant and Cell Physiology, 38(8): 945-951.
|
|
|
| [48] |
Sengupta S, Majumder A L. 2009. Insight into the salt tolerance factors of a wild halophytic rice, Porteresia coarctata: A physiological and proteomic approach. Planta, 229(4): 911-929.
doi: 10.1007/s00425-008-0878-y
pmid: 19130079
|
|
|
| [49] |
Shuyskaya E, Rakhmankulova Z, Voronin P, et al. 2015. Salt and osmotic stress tolerances of the C3-C4 xero-halophyte Bassia sedoides from two populations differ in productivity and genetic polymorphism. Acta Physiologiae Plantarum, 37: 236, doi: 10.1007/s11738-015-1981-x.
|
|
|
| [50] |
Shuyskaya E, Rakhmankulova Z, Prokofieva M, et al. 2022. Intensity and duration of salinity required to form adaptive response in C4 halophyte Kochia prostrata (L.) Shrad. Frontiers of Plant Sciences, 13(6): 955880, doi: 10.3389/fpls.2022.955880.
|
|
|
| [51] |
Shuyskaya E, Rakhmankulova Z, Prokofieva M, et al. 2023. Impact of salinity, elevated temperature, and their interaction with the photosynthetic efficiency of halophyte crop Chenopodium quinoa Willd. Agriculture, 13: 1198, doi: 10.3390/agriculture13061198.
|
|
|
| [52] |
Siadjeu C, Lauterbach M, Kadereit G. 2021. Insights into regulation of C2 and C4 photosynthesis in Amaranthaceae/ Chenopodiaceae using RNA-Seq. International Journal of Molecular Sciences, 22(22): 12120, doi: 10.3390/ijms222212120.
|
|
|
| [53] |
Soltis D E, Soltis P S. 1990. Isozymes in Plant Biology. London: Springer, 268.
|
|
|
| [54] |
Szabados L, Savouré A. 2010. Proline: A multifunctional amino acid. Trends in Plant Science, 15(2): 89-97.
doi: 10.1016/j.tplants.2009.11.009
pmid: 20036181
|
|
|
| [55] |
Voronin P Y. 2014. Experimental installation for measurements of chlorophyll fluorescence, CO2 exchange, and transpiration of a detached leaf. Russian Journal of Plant Physiology, 61: 269-273.
|
|
|
| [56] |
Voznesenskaya E V, Koteyeva N K, Akhani H, et al. 2013. Structural and physiological analyses in Salsoleae (Chenopodiaceae) indicate multiple transitions among C3, intermediate, and C4 photosynthesis. Journal of Experimental Botany, 64(12): 3583- 3604.
doi: 10.1093/jxb/ert191
pmid: 23881394
|
|
|
| [57] |
Wojnicka-Poltorak A, Chudzinska E, Shuiskay E, et al. 2002. Izoenzymatic and cytological studies of some Asiatic species of the genus Salsola. Acta Societatis Botanicorum Poloniae, 71(3): 115-120
|
|
|
| [58] |
Yamori W, Shikanai T. 2016. Physiological functions of cyclic electron transport around photosystem I in sustaining photosynthesis and plant growth. Annual Review of Plant Biology, 67: 81-106.
doi: 10.1146/annurev-arplant-043015-112002
pmid: 26927905
|
|
|
| [59] |
Yeh F C, Yang R C, Boyle T. 1999. POPGEN version 1.32 Microsoft windows-based freeware for populations genetic analysis. [2023-12-19]. https://api.semanticscholar.org/CorpusID:59539890.
|
|
|
| [60] |
Yi C, Yao K, Cai S, et al. 2015. High atmospheric carbon dioxide-dependent alleviation of salt stress is linked to respiratory burst oxidase 1 (RBOH1)-dependent H2O2 production in tomato (Solanum lycopersicum). Journal of Experimental Botany, 66(22): 7391-7404.
|
|
|
| [61] |
Yorimitsu Y, Kadosono A, Hatakeyama Y, et al. 2019. Transition from C3 to proto-Kranz to C3-C4 intermediate type in the genus Chenopodium (Chenopodiaceae). Journal of Plant Research, 132: 839-855.
|
|
|
| [62] |
Zandalinas S I, Fritschi F B, Mittler R. 2020. Signal transduction networks during stress combination. Journal of Experimental Botany, 71(5): 1734-1741.
doi: 10.1093/jxb/erz486
pmid: 31665392
|
|
|
| [63] |
Zandalinas S I, Mittler R. 2022. Plant responses to multifactorial stress combination. New Phytologist, 234(4): 1161-1167.
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|