| Research article |
|


|
|
|
| Spatiotemporal niche separation mechanisms of water utilization strategies in the desert steppe plant communities, northern China |
SONG Kechen1, HU Haiying1,*( ), ZHANG Hao1, GUAN Siyu1, DENG Wenhui1, YONG Jiayi1, ZHAO Xiaona1, WANG Xing2 ), ZHANG Hao1, GUAN Siyu1, DENG Wenhui1, YONG Jiayi1, ZHAO Xiaona1, WANG Xing2 |
1College of Forestry and Prataculture, Ningxia University, Yinchuan 750021, China
2Chengdu Institute of Biology, Chinese Academy of Science, Chengdu 610041, China |
|
|
|
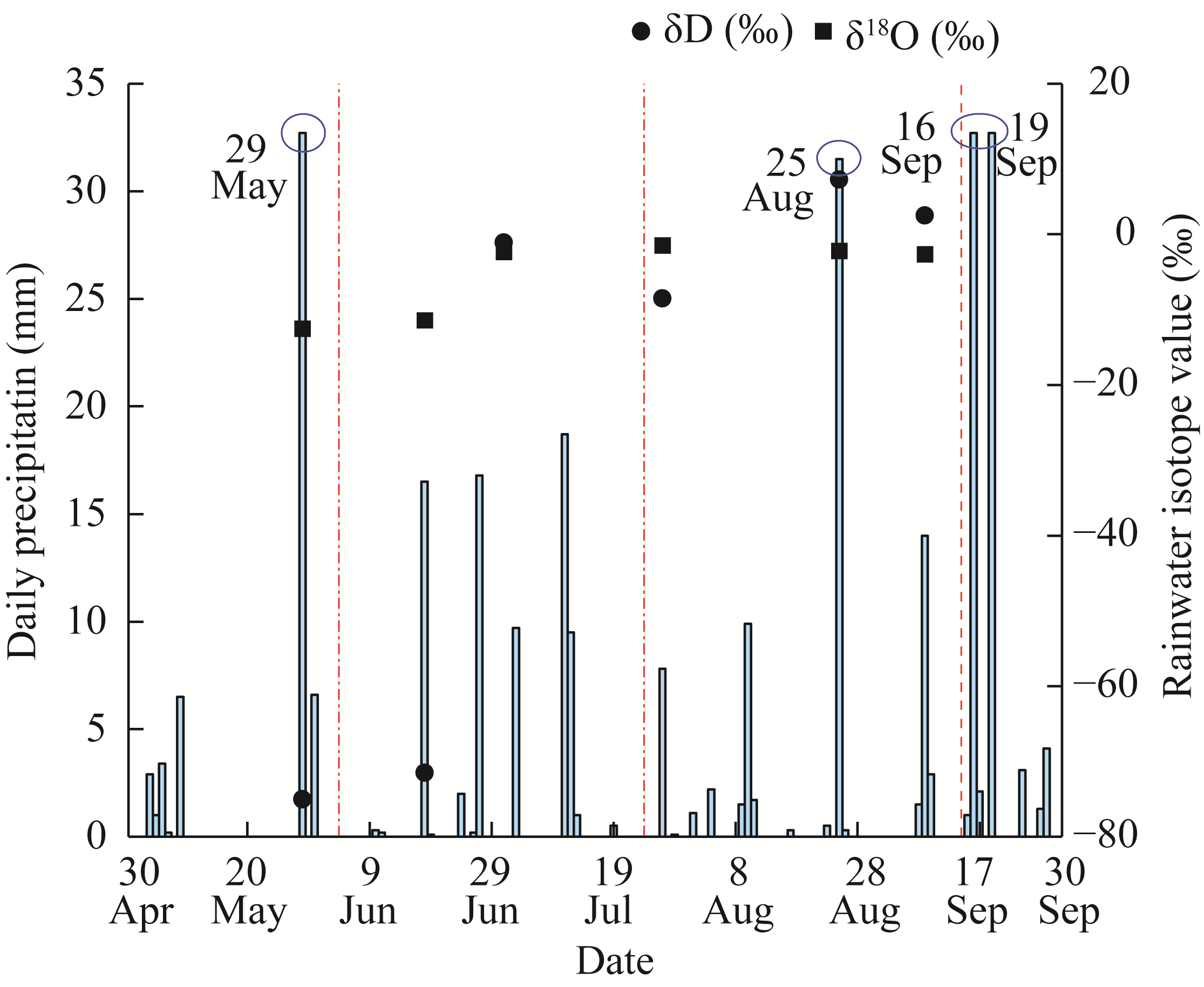
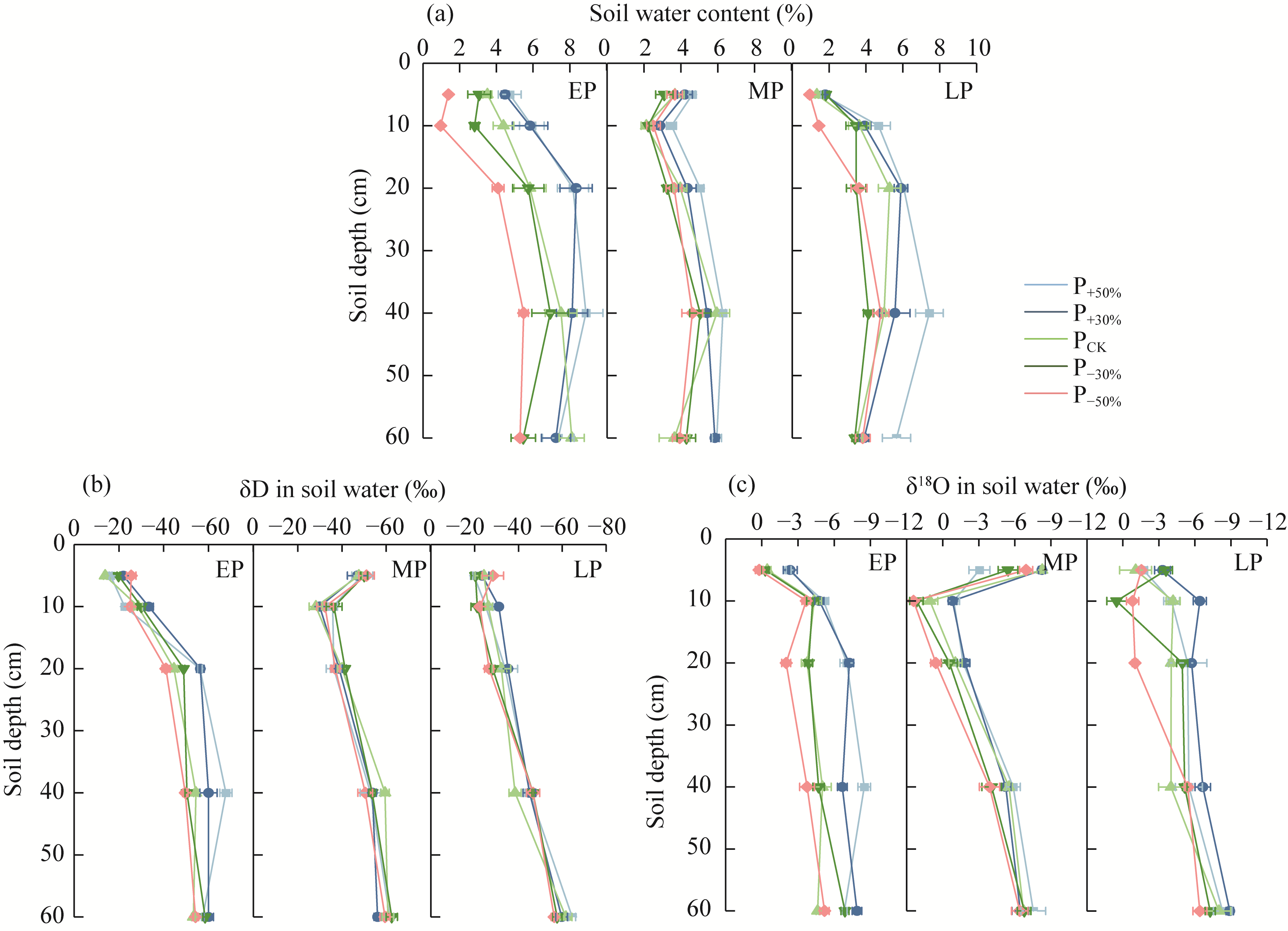
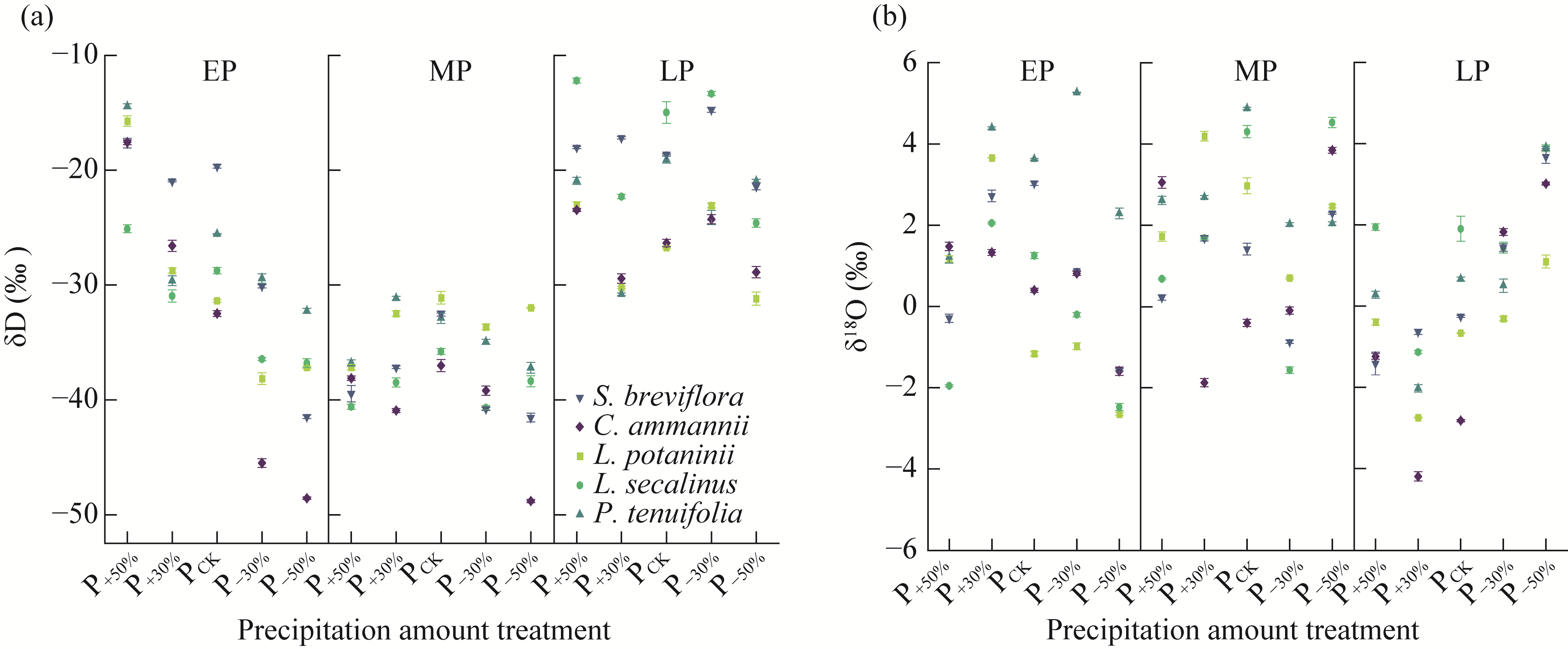
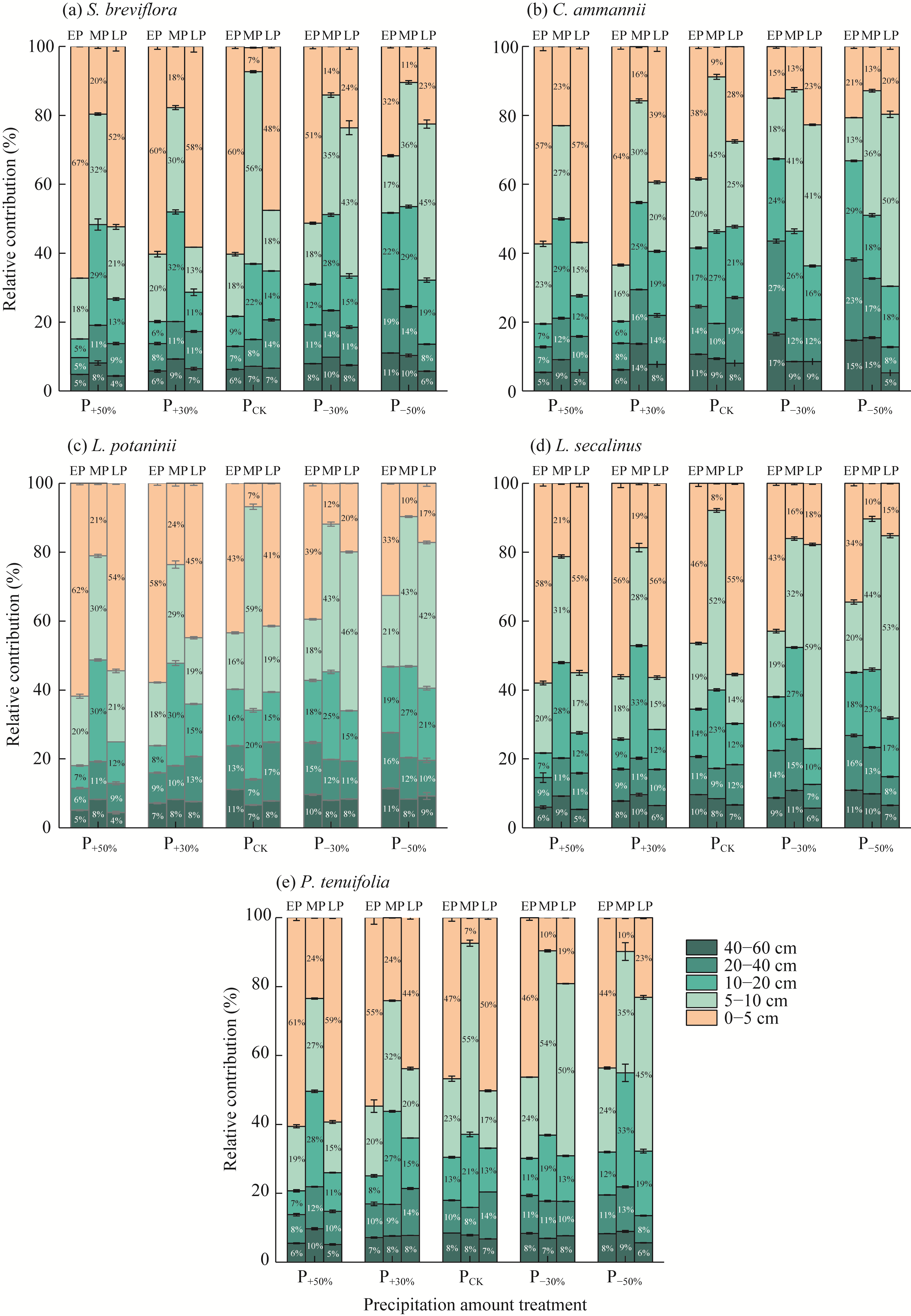
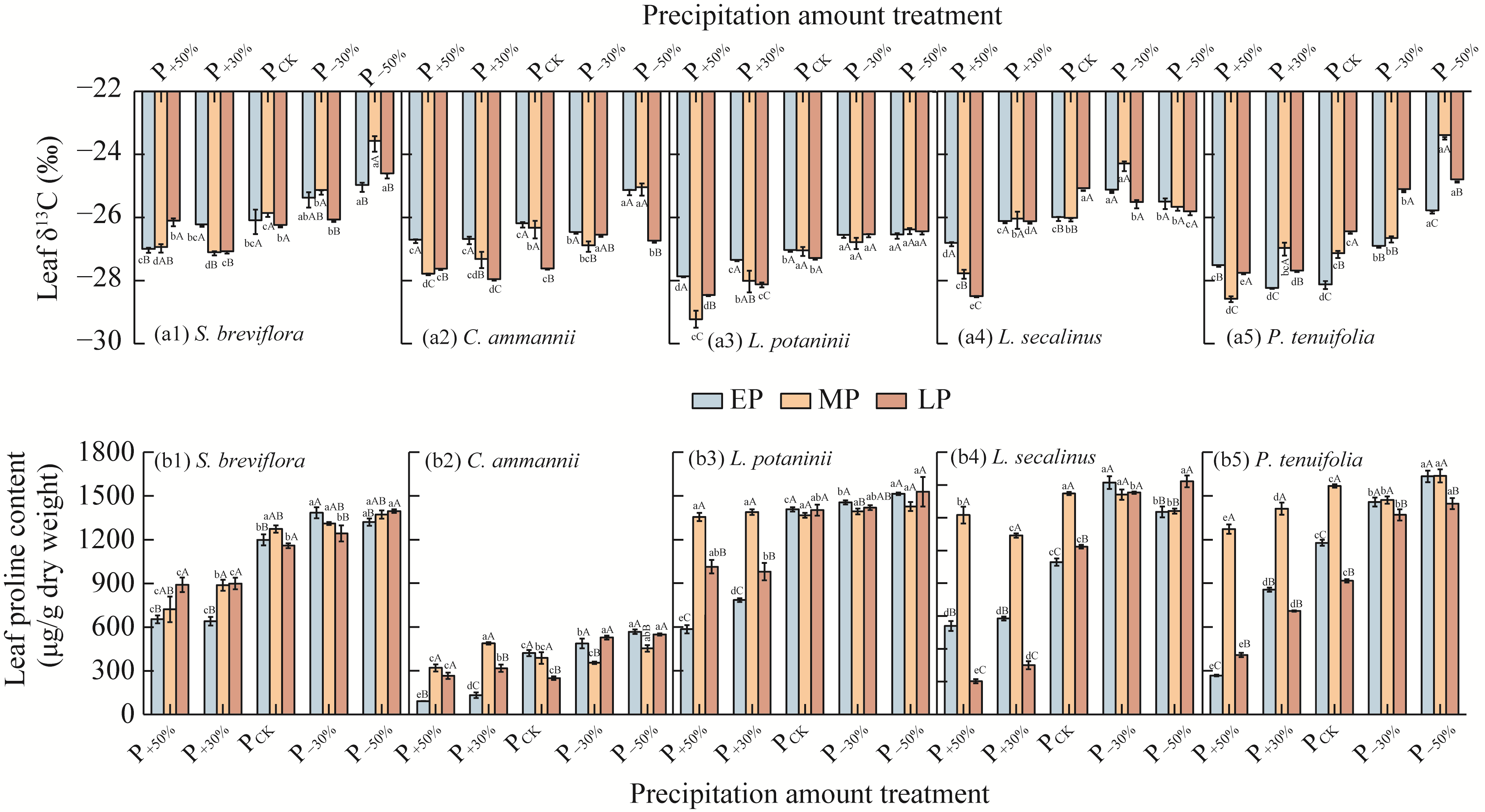
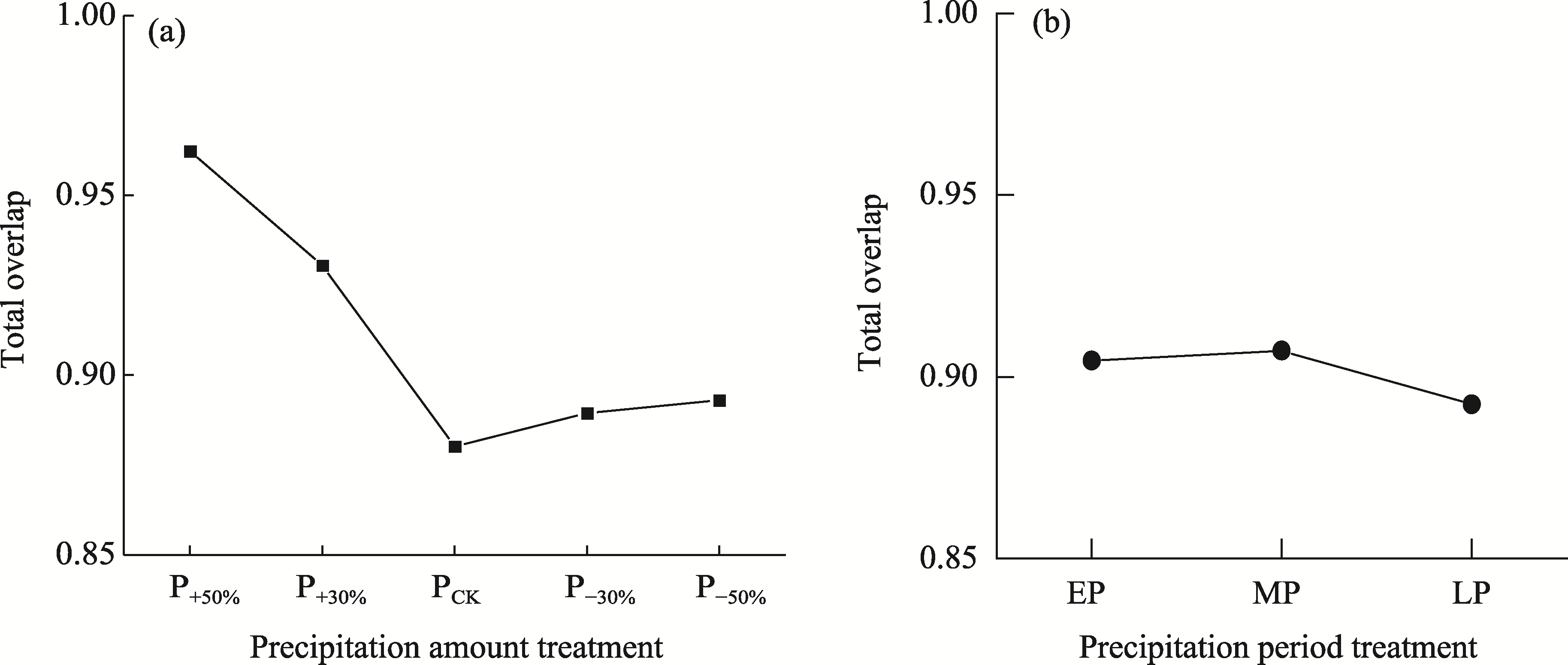
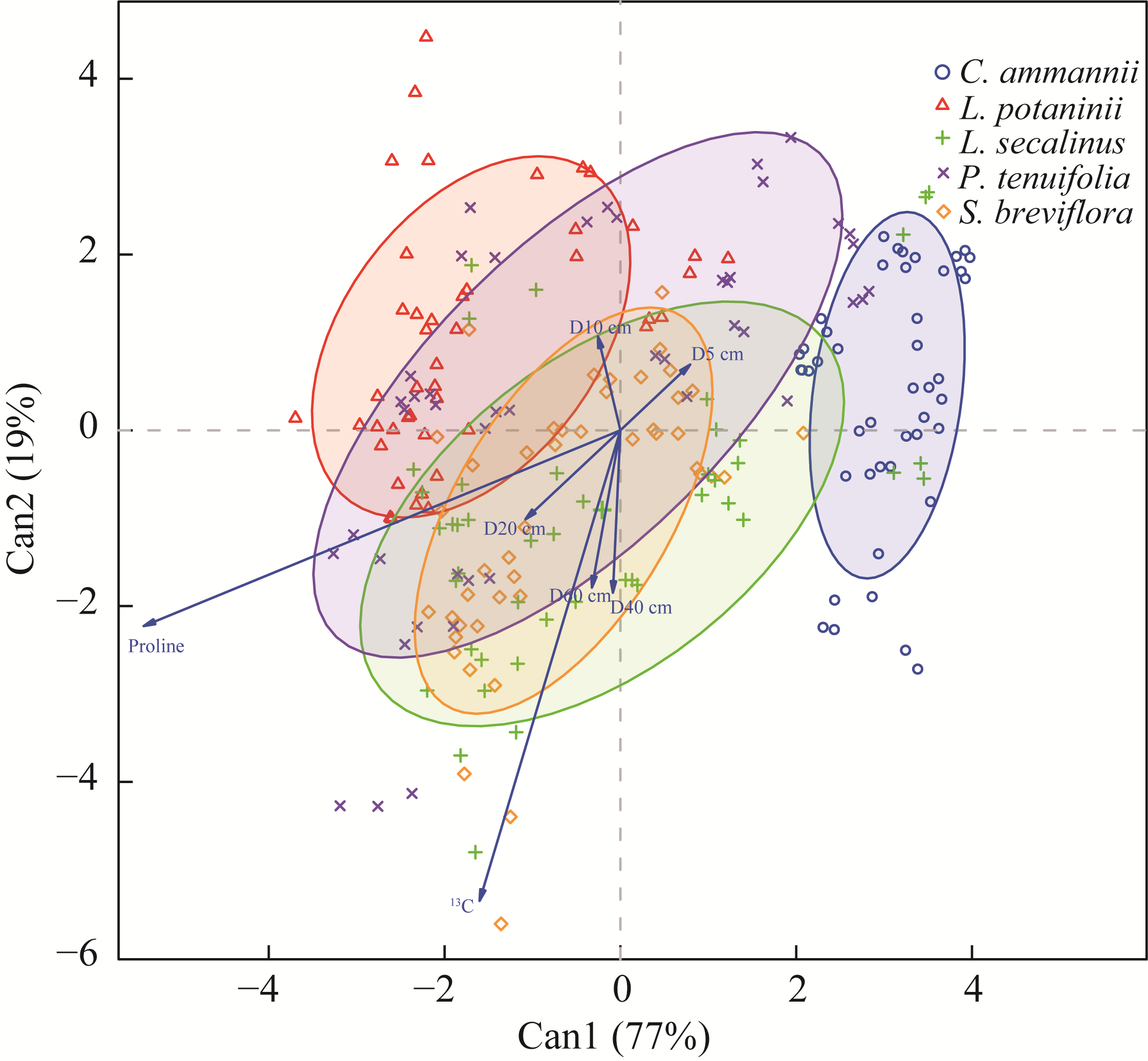
Abstract Amid global precipitation changes, it remains unclear whether hydrological niche separation (HNS) mechanisms apply to herbaceous plant communities in desert steppes are severely affected by seasonal drought. How these plants access limited water and tolerate drought to coexist also remains unverified. In this study, we employed stable isotope techniques to examine water acquisition and drought adaptation in coexisting species of the desert steppe in northern China under five precipitation treatments, i.e., decreased 50%, decreased 30%, ambient, increased 30%, and increased 50% precipitation. The following results showed that: (1) water sources of coexisting species shifted with changes in precipitation amount and timing, i.e., all coexisting plants exhibited preferential utilization of surface soil moisture. When surface soil moisture was scarce, they shifted to deeper water sources, and when deep water sources remained scarce, they were forced to compete more intensely for surface water sources; (2) community's HNS was affected by precipitation amount but not by timing, i.e., with adequate soil moisture, plant water source ranges expanded, reducing overlap and enhancing HNS, whereas under extreme drought, the range contracted and increased the overlap, although HNS remained stable; and (3) water acquisition strategies of coexisting species differed along hydrological niche axis defined by water stress adaptability (i.e., stable carbon isotope composition and proline content). Convolvulus ammannii Desr. had the strongest drought adaptation, although its strategy showed a weak correlation with water uptake. Stipa breviflora Griseb., with moderate drought resistance, adopted a water-conserving strategy that was suitable for extreme drought. Leymus secalinus (Georgi) Tzvelev, Polygala tenuifolia Willd., and Larix potaninii Batalin showed resource-dependent and flexible water strategies, thriving in wetter soils but struggling under extreme drought. Our findings indicated that herbaceous species in desert steppes adapted their water uptake and drought tolerance strategies according to changes in precipitation amount and timing. As a core regulatory mechanism, HNS (under increasing precipitation variability due to climate change) not only supports species coexistence by reducing interspecific competition, but also promotes efficient soil moisture use. This mechanism enhances community drought resistance and contributes to ecosystem stability. Overall, this study provides key ecological evidence for understanding plant community adaptation in arid and semi-arid areas facing the influence of global climate change.
|
|
Received: 23 April 2025
Published: 31 December 2025
|
|
Corresponding Authors:
*HU Haiying (E-mail: haiying@nxu.edu.cn)
|
| Cite this article:
SONG Kechen, HU Haiying, ZHANG Hao, GUAN Siyu, DENG Wenhui, YONG Jiayi, ZHAO Xiaona, WANG Xing. Spatiotemporal niche separation mechanisms of water utilization strategies in the desert steppe plant communities, northern China. Journal of Arid Land, 2025, 17(12): 1741-1760.
URL:
http://jal.xjegi.com/10.1007/s40333-025-0034-6 OR http://jal.xjegi.com/Y2025/V17/I12/1741
|
|
|
| [1] |
Anjum S A, Xie X Y, Wang L C, et al. 2011. Morphological, physiological and biochemical responses of plants to drought stress. African Journal of Agricultural Research, 6(9): 2026-2032.
|
|
|
| [2] |
Araya Y N, Silvertown J, Gowing D J, et al. 2011. A fundamental, ecohydrological basis for niche segregation in plant communities. New Phytologist, 189(1): 253-258.
doi: 10.1111/nph.2010.189.issue-1
|
|
|
| [3] |
Bachmann D, Gockele A, Ravenek JM, et al. 2015. No evidence of complementary water use along a plant species richness gradient in temperate experimental grasslands. PLoS ONE, 10(1): e0116367, doi:10.1371/journal.pone.0116367.
|
|
|
| [4] |
Bartholomeus R P, Witte J P M, Bodegom P M V, et al. 2011. Climate change threatens endangered plant species by stronger and interacting water-related stresses. Journal of Geophysical Research: Biogeosciences, 116(G4): G04023, doi:10.1029/2011JG001693.
|
|
|
| [5] |
Beyer M K, Kühnhammer K, Dubbert M. 2020. In situ measurements of soil and plant water isotopes: A review of approaches, practical considerations and a vision for the future. Hydrology and Earth System Sciences, 24(9): 4413-4440.
doi: 10.5194/hess-24-4413-2020
|
|
|
| [6] |
Bimler M D, Stouffer D B, Lai H R, et al. 2018. Accurate predictions of coexistence in natural systems require the inclusion of facilitative interactions and environmental dependency. Journal of Ecology, 106(5): 1839-1852.
doi: 10.1111/jec.2018.106.issue-5
|
|
|
| [7] |
Blanco-Sánchez M, Ramos-Muñoz M, Pías B, et al. 2022. Natural selection favours drought escape and an acquisitive resource-use strategy in semi-arid Mediterranean shrubs. Functional Ecology, 36(9): 2289-2302.
doi: 10.1111/fec.v36.9
|
|
|
| [8] |
Braun L, Kadmon R, Tomiolo S, et al. 2022. Is more less? A comprehensive experimental test of soil depth effects on grassland diversity. Oikos, 2022(5): e08535, doi:10.1111/oik.08535.
|
|
|
| [9] |
Chen F L, Wang S J, Wu X X, et al. 2021. Local meteoric water lines in a semi-arid setting of Northwest China using multiple methods. Water, 13(17): 2380, doi:10.3390/w13172380.
|
|
|
| [10] |
Chen J, Xu Q, Gao D Q, et al. 2017. Differential water use strategies among selected rare and endangered species in West Ordos Desert of China. Journal of Plant Ecology, 10(4): 660-669.
doi: 10.1093/jpe/rtw059
|
|
|
| [11] |
Cheng X L, An S Q, Li B, et al. 2006. Summer rain pulse size and rainwater uptake by three dominant desert plants in a desertified grassland ecosystem in northwestern China. Plant Ecology, 184: 1-12.
doi: 10.1007/s11258-005-9047-6
|
|
|
| [12] |
Chesson P. 2000. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 31(1): 343-366.
doi: 10.1146/ecolsys.2000.31.issue-1
|
|
|
| [13] |
Detto M, Montaldo N, Albertson J D, et al. 2006. Soil moisture and vegetation controls on evapotranspiration in a heterogeneous Mediterranean ecosystem on Sardinia, Italy. Water Resources Research, 42(8): W08419, doi:10.1029/2005WR004693.
|
|
|
| [14] |
Dwyer C, Pakeman RJ, Jones L, et al. 2021. Fine-scale hydrological niche segregation in coastal dune slacks. Journal of Vegetation Science, 32(5): e13085, doi:10.1111/jvs.13085.
|
|
|
| [15] |
Ehleringer J R, Dawson T E. 1992. Water uptake by plants: perspectives from stable isotope composition. Plant, Cell and Environment, 15(9): 1073-1082.
doi: 10.1111/pce.1992.15.issue-9
|
|
|
| [16] |
Fargione J, Tilman D. 2005. Niche differences in phenology and rooting depth promote coexistence with a dominant C4 bunchgrass. Oecologia, 143(4): 598-606.
pmid: 15791430
|
|
|
| [17] |
Fort F, Volaire F, Guilioni L, et al. 2017. Root traits are related to plant water-use among rangeland Mediterranean species. Functional Ecology, 31(9): 1700-1709.
doi: 10.1111/fec.2017.31.issue-9
|
|
|
| [18] |
Garrett R H, Grisham C M. 1999. Biochemistry (2nd ed). Fort Worth: Harcourt Inc.
|
|
|
| [19] |
Grime J P. 1997. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist, 111(982): 1169-1194.
doi: 10.1086/283244
|
|
|
| [20] |
Gross N, Bagousse-Pinguet Y L, Liancourt P, et al. 2025. Unveiling ecological assembly rules from commonalities in trait distributions. Ecology Letters, 24(8): 1668-1680.
doi: 10.1111/ele.v24.8
|
|
|
| [21] |
Guderle M, Bachmannd D, Milcu A, et al. 2018. Dynamic niche partitioning in root water uptake facilitates efficient water use in more diverse grassland plant communities. Function Ecology, 32(1): 214-227.
doi: 10.1111/fec.2018.32.issue-1
|
|
|
| [22] |
Gui J, Li Z X, Du F, et al. 2024. Vegetation restoration strategies based on plant water use patterns. Science of the Total Environment, 924: 171611, doi:10.1016/j.scitotenv.2024.171611.
|
|
|
| [23] |
Guo J S, Hungate B A, Kolb T E, et al. 2018. Water source niche overlap increases with site moisture availability in woody perennials. Plant Ecology, 219(6): 719-735.
doi: 10.1007/s11258-018-0829-z
|
|
|
| [24] |
Harpole W. 2010. Neutral theory of species diversity. Nature Education Knowledge, 3(10): 60.
|
|
|
| [25] |
Hoekstra N J, Finn J A, Lüscher A. 2014. The effect of drought and interspecific interactions on depth of water uptake in deep- and shallow-rooting grassland species as determined by δ18O natural abundance. Biogeosciences, 11(16): 4493-4506.
doi: 10.5194/bg-11-4493-2014
|
|
|
| [26] |
Holdo R M, Nippert J B. 2023. Linking resource- and disturbance-based models to explain tree-grass coexistence in savannas. New Phytologist, 237(6): 1966-1979.
doi: 10.1111/nph.v237.6
|
|
|
| [27] |
Horton J L, Hart S C, Kolb T E. 2003. Physiological condition and water source use of Sonoran Desert riparian trees at the Bill Williams River, Arizona, USA. Isotopes in Environmental and Health Studies, 39(1): 69-82.
pmid: 12812256
|
|
|
| [28] |
Hu H Y, Zhu L, Li H X, et al. 2021. Seasonal changes in the water-use strategies of three herbaceous species in a native desert steppe of Ningxia, China. Journal of Arid Land, 13(2): 109-122.
doi: 10.1007/s40333-021-0051-z
|
|
|
| [29] |
Ivanov V Y, Hutyra L R, Wofsy S C, et al. 2012. Root niche separation can explain avoidance of seasonal drought stress and vulnerability of overstory trees to extended drought in a mature Amazonian forest. Water Resources Research, 48(12): WR011972, doi:10.1029/2012WR011972.
|
|
|
| [30] |
Kang J, Peng Y F, Xu W F. 2022. Crop root responses to drought stress: molecular mechanisms, nutrient regulations, and interactions with microorganisms in the rhizosphere. International Journal of Molecular Sciences, 23(16): 9310, doi:10.3390/ijms23169310.
|
|
|
| [31] |
Kannenberg S A, Guo J S, Novick K A, et al. 2022. Opportunities, challenges and pitfalls in characterizing plant water-use strategies. Functional Ecology, 36(1): 24-37.
doi: 10.1111/fec.v36.1
|
|
|
| [32] |
Kulmatiski A, Adler P B, Foley K M. 2020a. Hydrologic niches explain species coexistence and abundance in a shrub-steppe system. Journal of Ecology, 108(3): 998-1008.
doi: 10.1111/jec.v108.3
|
|
|
| [33] |
Kulmatiski A, Beard M K H, Holdrege C, et al. 2020b. Small differences in root distributions allow resource niche partitioning. Ecology and Evolution, 10(18): 9776-9787.
doi: 10.1002/ece3.v10.18
|
|
|
| [34] |
Kulmatiski A, Beard K H. 2022. A modern two-layer hypothesis helps resolve the savanna problem. Ecology Letters. 25(9): 1952-1960.
doi: 10.1111/ele.14067
pmid: 35834518
|
|
|
| [35] |
Li B B, Zang M J, Che C W, et al. 2024. Water utilization strategy of Ziziphus jujuba under different sand cover thicknesses based on stable isotope tracing. Chinese Journal of Plant Ecology, 48(9): 1202-1212. (in Chinese)
doi: 10.17521/cjpe.2023.0295
|
|
|
| [36] |
Michalet R, Delerue F, Liancourt P, et al. 2021. Are complementarity effects of species richness on productivity the strongest in species-rich communities? Journal of Ecology, 109(5): 2038-2046.
doi: 10.1111/jec.v109.5
|
|
|
| [37] |
Mueller K E, Tilman D, Fornara D A, et al. 2013. Root depth distribution and the diversity-productivity relationship in a long-term grassland experiment. Ecology, 94(4): 787-793.
doi: 10.1890/12-1399.1
|
|
|
| [38] |
Nippert J B, Knapp A K. 2007. Soil water partitioning contributes to species coexistence in tallgrass prairie. Oikos, 116(6): 1017-1029.
doi: 10.1111/oik.2007.116.issue-6
|
|
|
| [39] |
Oram N J, Ravenek J M, Barry K E, et al. 2018. Below-ground complementarity effects in a grassland biodiversity experiment are related to deep-rooting species. Journal of Ecology, 106(1): 265-277.
doi: 10.1111/jec.2018.106.issue-1
|
|
|
| [40] |
Pastore M. 2018. Overlapping: A R package for estimating overlapping in empirical distributions. The Journal of Open Source Software, 3(32): 1023, doi:10.21105/joss.01023.
|
|
|
| [41] |
Prechsl U E, Burri S, Gilgen A K, et al. 2015. No shift to a deeper water uptake depth in response to summer drought of two lowland and sub-alpine C3-grasslands in Switzerland. Oecologia, 177(1): 97-111.
doi: 10.1007/s00442-014-3092-6
pmid: 25273953
|
|
|
| [42] |
Sala O E, Chapin F S, Armesto J J, et al. 2000. Global biodiversity scenarios for the year 2100. Science, 287(5459): 1770-1774.
doi: 10.1126/science.287.5459.1770
pmid: 10710299
|
|
|
| [43] |
Schlaepfer D R, Bradford J B, Lauenroth W K, et al. 2017. Climate change reduces extent of temperate drylands and intensifies drought in deep soils. Nature Communications, 8: 14196, doi:10.1038/ncomms14196.
pmid: 28139649
|
|
|
| [44] |
Silvertown J, Dodd M E, Gowing D J G, et al. 1999. Hydrologically defined niches reveal a basis for species richness in plant communities. Nature, 400: 61-63.
doi: 10.1038/21877
|
|
|
| [45] |
Silvertown J. 2004. Plant coexistence and the niche. Trends in Ecology and Evolution, 19: 605-611.
doi: 10.1016/j.tree.2004.09.003
|
|
|
| [46] |
Silvertown J, Araya Y, Gowing D. 2015. Hydrological niches in terrestrial plant communities: A review. Journal of Ecology, 103(1): 93-108.
doi: 10.1111/jec.2015.103.issue-1
|
|
|
| [47] |
Stock B C, Semmens B X. 2016. Unifying error structures in commonly used biotracer mixing models. Ecology, 97(10): 2562-2569.
doi: 10.1002/ecy.1517
pmid: 27859126
|
|
|
| [48] |
Swanson H, Lysy M, Power M, et al. 2015. A new probabilistic method for quantifying n-dimensional ecological niches and niche overlap. Ecology, 96(2): 318-324.
pmid: 26240852
|
|
|
| [49] |
Wainwright C E, HilleRisLambers J, Lai H R, et al. 2018. Distinct responses of niche and fitness differences to water availability underlie variable coexistence outcomes in semi-arid annual plant communities. Journal of Ecology, 107(1): 293-306.
doi: 10.1111/jec.2019.107.issue-1
|
|
|
| [50] |
Wang J, Fu B J, Lü N, et al. 2017. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semiarid Loess Plateau. Science of the Total Environment, 609(1): 27-37.
doi: 10.1016/j.scitotenv.2017.07.133
|
|
|
| [51] |
Wang Y T, Shen Y J, Xie Y Z, et al. 2023. Changes in precipitation have both direct and indirect effects on typical steppe aboveground net primary productivity in Loess Plateau, China. Plant and Soil, 484(1-2): 503-515.
doi: 10.1007/s11104-022-05812-8
|
|
|
| [52] |
Weides S E, Hájek T, Liancourt P, et al. 2024. Belowground niche partitioning is maintained under extreme drought. Ecology, 105(1): e4198, doi:10.1002/ecy.4198.
|
|
|
| [53] |
Williams D G, Ehleringer J R. 2000. Intra and interspecific variation for summer precipitation use in pinyon-juniper woodlands. Ecological Monographs, 70: 517-537.
|
|
|
| [54] |
Wu J E, Zeng H H, Zhao F, et al. 2022. Plant hydrological niches become narrow but stable as the complexity of interspecific competition increases. Agricultural and Forest Meteorology, 320: 108953, doi:10.1016/j.agrformet.2022.108953.
|
|
|
| [55] |
Xing B B, An H, Liu S X, et al. 2023. Effects of rainfall change on plant community biomass and its trade-off in desert grassland. Acta Agrestia Sinica, 31(10): 3103-3113. (in Chinese)
|
|
|
| [56] |
Zhang B, Zhu J J, Liu H M, et al. 2014. Effects of extreme rainfall and drought events on grassland ecosystems. Chinese Journal of Plant Ecology, 38(9): 1008-1018. (in Chinese)
doi: 10.3724/SP.J.1258.2014.00095
|
|
|
| [57] |
Zhang H, Song K C, Hu H Y, et al. 2024. Variability in precipitation influences the water sourcing and adaptive strategies of key plant species within the desert steppe ecosystem. Ecological Indicators, 158: 111333, doi:10.1016/j.ecolind.2023.111333.
|
|
|
| [58] |
Zhou H, Zhao W Z, He Z B. 2017. Water sources of Nitraria sibirica and response to precipitation in two desert habitats. Chinese Journal of Applied Ecology, 28(7): 2083-2092. (in Chinese)
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|