Seasonal changes in the water-use strategies of three herbaceous species in a native desert steppe of Ningxia, China
HU Haiying1,2,3, ZHU Lin1,2, LI Huixia3, XU Dongmei3, XIE Yingzhong1,2,3,*()
1Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration of North-western China, Ningxia University, Yinchuan 750021, China 2Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in North-western China of Ministry of Education, Ningxia University, Yinchuan 750021, China 3School of Agriculture, Ningxia University, Yinchuan 750021, China
Frequent periods of drought conditions are known to limit plant performance, primary production, and ecosystem stability in arid and semi-arid desert steppe environments. Plants often avoid competition by shifting their water use seasonally, which affects the water-use patterns of dominant species as well as the composition and structure of plant communities. However, the water-use strategies of dominant herbaceous species, which grow under natural field conditions in the desert steppe region of Ningxia Hui Autonomous Region, China, are poorly known. Here, we explored the possible sources of water uptake and water-use efficiency (WUE) of three dominant herbaceous plant species (Stipa breviflora, Agropyron mongolicum, and Glycyrrhiza uralensis) in a native desert steppe in the semi-arid area of Ningxia through an analysis of multiple parameters, including (1) the stable isotopic oxygen and hydrogen (δ 18O and δ 2H) compositions of precipitation, soil water, and stem water, (2) the carbon isotope ( 13C) composition of leaves, and (3) the soil water contents, based on field sampling across varying water conditions from June to September, 2017. Frequent small precipitation events replenished shallow soil water, whereas large events only percolated down to the deep soil layers. Changes in soil water availability affected the water-use patterns of plants. Generally, during light precipitation periods, the deep root system of G. uralensis accessed deeper (>80 cm) soil water, whereas S. breviflora and A. mongolicum, which only have shallow roots, primarily absorbed water from the shallow and middle soil layers. As precipitation increased, all three plant species primarily obtained water from the shallow soil layers. Variation in soil water uptake between the dry and wet seasons enabled plants to make better use of existing satoil water. In addition, the δ 13C values of G. uralensis and S. breviflora were higher than those of A. mongolicum. The δ 13C values of the three plant species were significantly negatively correlated with soil water content. Therefore, G. uralensis and S. breviflora maintained a higher WUE through their conservative and water-saving strategies across the entire growing season. In contrast, A. mongolicum, with a relatively low WUE in the wet season but a high WUE in the dry season, exhibited a more flexible water-use strategy. The different water-use strategies of these dominant plant species demonstrated the mechanisms by which plant communities can respond to drought.
Received: 26 March 2020
Published: 10 February 2021
HU Haiying, ZHU Lin, LI Huixia, XU Dongmei, XIE Yingzhong. Seasonal changes in the water-use strategies of three herbaceous species in a native desert steppe of Ningxia, China. Journal of Arid Land, 2021, 13(2): 109-122.
Fig. 1Geographic location of the study area (a) and photographs of Stipa breviflora (b), Agropyron mongolicum (c), and Glycyrrhiza uralensis (d)
Fig. 2Photographs showing compacted sierozem in the form of unconsolidated sediment in the A. mongolicum site (a) and compacted aeolian sandy soil in the S. breviflora+G. uralensis site (b)
Species
Family
Height (cm)
Density (plants/m2)
Coverage (%)
Roots
Agropyron mongolicum
Gramineae
28.13±12.5
31.40±11.70
12.80±5.40
Sparse clump-type, with the rooting depth of 80-100 cm
Stipa breviflora
Gramineae
12.93±2.30
13.40±5.60
21.60±13.50
Dense bundle-type, with the rooting depth of 60-80 cm
Glycyrrhiza uralensis
Leguminosae
21.60±5.10
8.80±3.60
5.20±2.90
Rhizome-type, with the rooting depth of 150-200 cm
Table 1 Characteristics of the dominant plant species
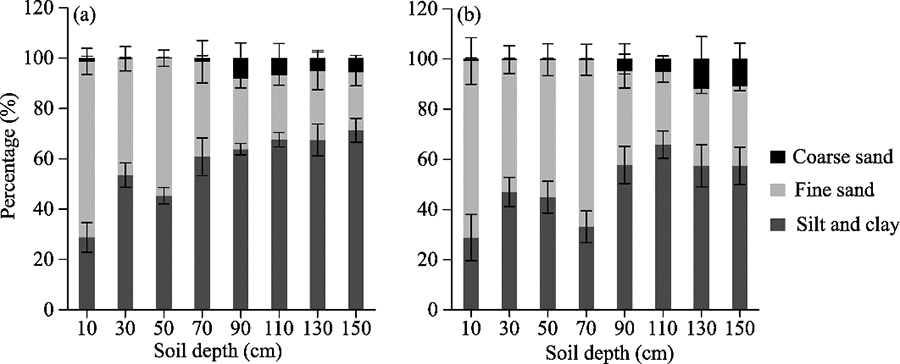
Fig. 3Variations in soil texture as a function of depth in A. mongolicum (a) and S. breviflora+G. uralensis (b) sites. Error bar represents standard error of the mean of three observations. The particle size classes were defined as coarse sand (0.10-2.00 mm), fine sand (0.05-0.10 mm), and silt and clay (<0.05 mm).
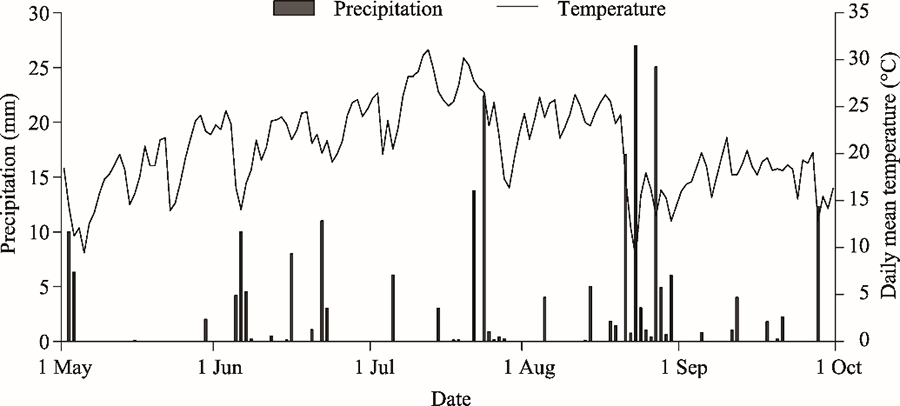
Fig. 4Daily precipitation and temperature from May to October 2017
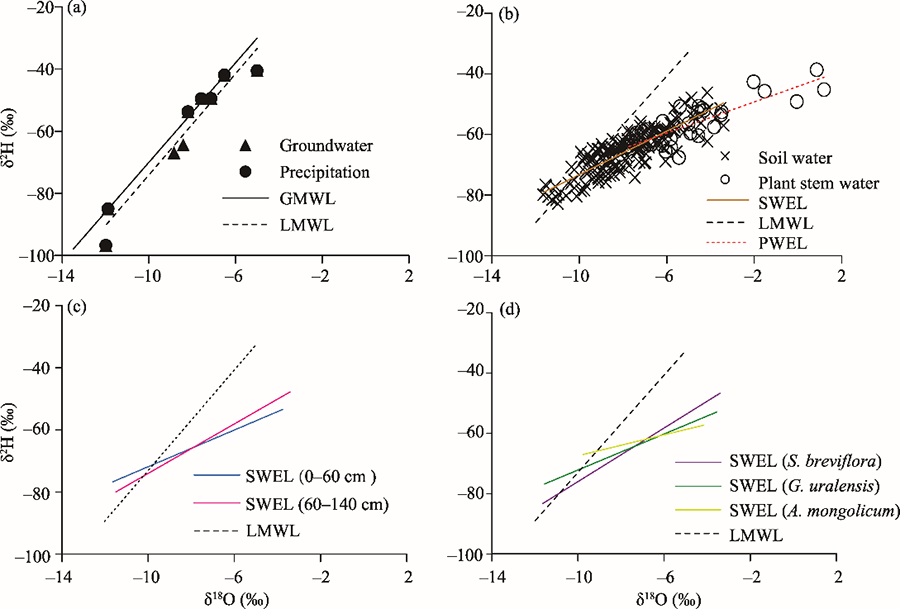
Fig. 5Curves of δ2H and δ18O values of different water sources. (a), precipitation and groundwater (GW); (b), plant stem water and soil water; (c), soil water of 0-60 and 60-140 cm layers; (d), soil water of plant communities. GMWL, global meteoric water line (δ2H=8δ18O+10); LMWL, local meteoric water line (δ2H=8.10δ18O+7.18; R2=0.9332; n=9); SWEL, soil water evaporation line; PWEL, plant stem water evaporation line. The corresponding curve equations of SWEL and PWEL are shown in Table 2.
Item
Equation
Confidence interval of model parameters
R2
F value
df
Sig.
At 2.50% level
At 97.50% level
δ2H
δ18O
δ2H
δ18O
SWEL
δ2H=3.59δ18O-37.35
-39.94
3.27
-34.77
3.92
0.74
481.80
169
***
PWEL
δ2H=2.57δ18O-44.11
-47.88
1.82
-40.34
3.32
0.65
50.25
25
***
SWEL (0-60 cm)
δ2H=2.85δ18O-42.99
-47.48
2.27
-38.50
3.44
0.58
95.31
70
***
SWEL (60-140 cm)
δ2H=3.98δ18O-34.20
-37.27
3.61
-31.12
4.35
0.82
446.10
97
***
SWEL (A. mongolicum)
δ2H=4.45δ18O-31.57
-35.37
3.98
-27.77
4.91
0.86
364.80
59
***
SWEL (S. breviflora)
δ2H=2.99δ18O-42.33
-45.53
2.58
-39.12
3.38
0.77
220.10
64
***
SWEL (G. uralensis)
δ2H=1.74δ18O-50.07
-55.84
1.00
-44.30
2.47
0.35
22.71
43
***
Table 2 Confidence intervals and statistics of the curve equations of SWEL and PWEL
Factor
Item
Soil water content (%)
Soil water
δ2H value (‰)
δ18O value (‰)
Season
Early dry season
5.29c
-61.42a
-6.87a
Late dry season
6.34b
-66.77b
-8.20b
Wet season
9.05a
-68.43b
-8.32b
Sig.
***
***
***
Community
A. mongolicum
9.86a
-66.49a
-7.80a
S. breviflora
5.78b
-65.58a
-7.72a
G. uralensis
5.47b
-65.04a
-7.98a
Sig.
***
ns
ns
Soil layer
0-20 cm
4.80c
-63.22a
-7.06a
20-40 cm
7.14b
-64.34ab
-7.52ab
40-60 cm
5.17c
-65.93b
-8.08bc
60-80 cm
4.91c
-65.51ab
-7.93bc
80-100 cm
8.96a
-66.88bc
-8.31c
100-120 cm
8.91a
-69.18c
-8.59c
120-140 cm
9.17a
-64.99ab
-7.37ab
Sig.
***
***
***
Table 3 Comparisons in soil water content and the δ2H and δ18O values of soil water in different soil layers of different plant communities in different seasons
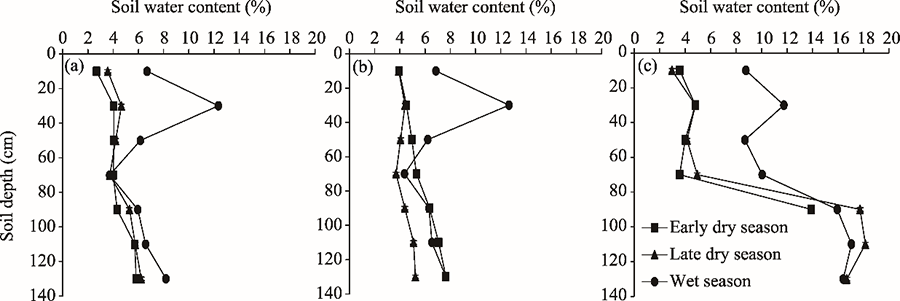
Fig. 6Soil water contents at 0-140 cm soil depths for S. breviflora (a), G. uralensis (b), and A. mongolicum (c) communities measured in different seasons
Factor
Item
δ13C value (‰)
δ2H value (‰)
δ18O value (‰)
Season
Early dry season
-26.23a
-60.69b
-5.04b
Late dry season
-26.07a
-49.43a
-2.72a
Wet season
-26.90b
-56.75b
-5.68b
Sig.
***
***
***
Plant species
A. mongolicum
-27.58b
-50.12a
-2.49a
S. breviflora
-25.85a
-56.81b
-4.96b
G. uralensis
-25.76a
-59.93b
-5.99b
Sig.
***
***
***
Season×Species
Sig.
***
***
***
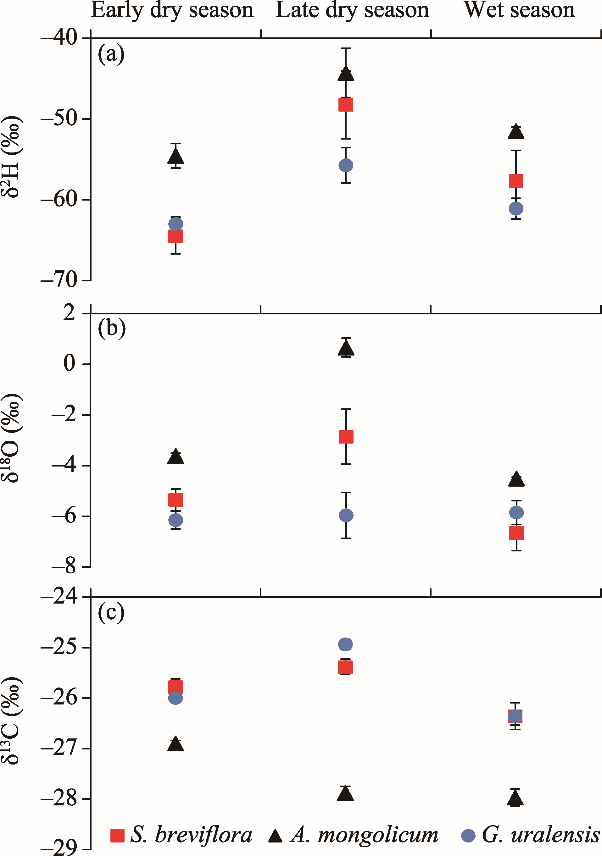
Table 4 Comparisons of the δ2H and δ18O values of stem water and δ13C values of leaves among different plant species in different seasons
Fig. 7δ2H (a) and δ18O (b) values of stem water and δ13C values (c) of leaves for S. breviflora, A. mongolicum, and G. uralensis in different seasons. Error bar represents standard error of mean of the observed values.
Species
Parameter
δ18O value
δ2H value
δ13C value
A. mongolicum
Soil water content
-0.00
0.37
-0.80**
δ18O value
1.00
0.79*
-0.26
δ2H value
0.79*
1.00
-0.46
δ13C value
-0.26
-0.46
1.00
S. breviflora
Soil water content
-0.50
-0.02
-0.77*
δ18O value
1.00
0.78*
0.89**
δ2H value
0.78*
1.00
0.55
δ13C value
0.89**
0.55
1.00
G. uralensis
Soil water content
0.16
-0.53
-0.93**
δ18O value
1.00
0.51
-0.18
δ2H value
0.51
1.00
0.65
δ13C value
-0.18
0.65
1.00
Table 5 Correlations among soil water content, δ2H and δ18O values of stem water, and δ13C values of leaves for A. mongolicum, S. breviflora, and G. uralensis communities
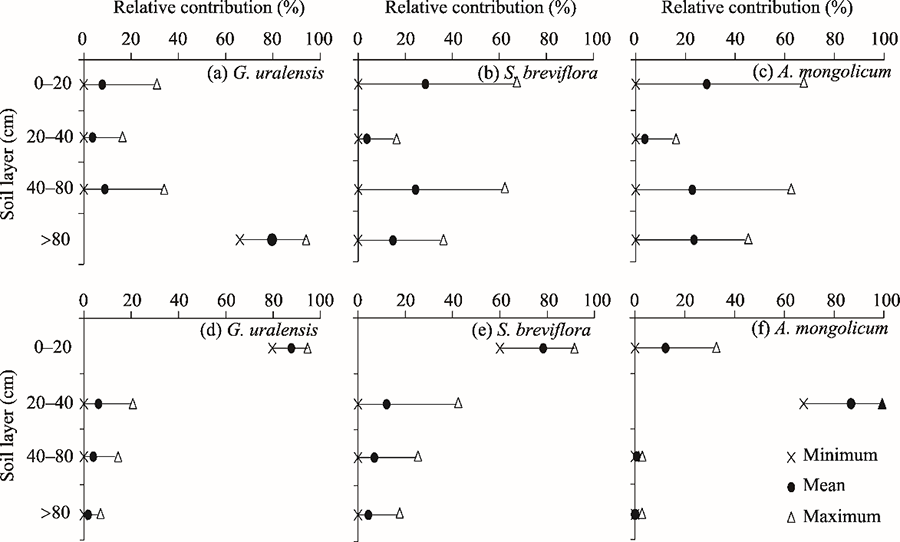
Fig. 8Relative contributions of potential water sources for G. uralensis, S. breviflora, and A. mongolicum in the different soil layers in the early dry season (a, b, c) and wet season (d, e, f). Bars represent the possible relative contribution ranges of potential water sources.
[1]
Altieri S, Mereu S, Cherubini P, et al. 2015. Tree-ring carbon and oxygen isotopes indicate different water use strategies in three Mediterranean shrubs at Capo Caccia. Sardinia, Italy. Trees Structure and Function, 5:1593-1603.
[2]
Antunes C, Díaz-Barradas M C, Zunzunegui M, et al. 2018. Water source partitioning among plant functional types in a semi-arid dune ecosystem. Journal of Vegetation Science, 29:671-683.
[3]
Araya Y N, Silvertown J, Gowing D J, et al. 2011. A fundamental, eco-hydrological basis for niche segregation in plant communities. New Phytologist, 189:253-258.
[4]
Auerswald K, Landinger C, Wittmer M, et al. 2010. 13Carbon allocated to the leaf growth zone of Poa pratensis reflects soil water and vapor pressure deficit. In: Schnyder H, Isselstein J, Taube F, et al. Grassland in a Changing World. Gottingen: Universität Göttingen Press, 857-859.
[5]
Bai Y F, Han X G, Wu J G, et al. 2004. Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature, 431:181-184.
pmid: 15356630
[6]
Bian Y Y, Song N P, Wang X, et al. 2015. Soil water deficit under different land-use type in desert steppe. Journal of Soil and Water Conservation, 29(1):201-206, 213. (in Chinese)
[7]
Bowen G J, Putnam A, Brooks J R, et al. 2018. Inferring the source of evaporated waters using stable H and O isotopes. Oecologia, 187:1025-1039.
doi: 10.1007/s00442-018-4192-5
pmid: 29955987
[8]
Burgess S S O, Adams M A, Turner N C, et al. 2000. Characterisation of hydrogen isotope profiles in an agroforestry system: implications for tracing water sources of trees. Agricultural Water Management, 3:229-241.
[9]
Chen G, Auerswald K, Schnyder H. 2016. 2H and 18O depletion of water close to organic surfaces . Biogeosciences, 13:3175-3186.
[10]
Chen J, Chen L, Song N P, et al. 2018. Soil in filtration characteristics of different soils types in desert steppe. Journal of Soil and Water Conservation, 32(4):18-23. (in Chinese)
[11]
Cheng X R, Huang M B, Shao M G, et al. 2008. Root distribution and soil water dynamics of Medicago sativa L. and Stipa breviflora Griseb. Acta Prataculturae Sinica, 2:170-175. (in Chinese)
[12]
Craig H. 1961. Isotopic variations in meteoric waters. Science, 3465:1702-1703.
[13]
Dai Y, Zheng X J, Tang L S, et al. 2015. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Plant and Soil, 389(1-2):73-87.
[14]
Dawson T E, Mambelli S, Plamboeck A H, et al. 2002. Stable isotopes in plant ecology. Annual Review of Ecology and Systematics, 33(1):507-559.
[15]
Deng W P, Zhang J, Zhang Z J, et al. 2017. Stable hydrogen and oxygen isotope compositions in soil-plant-atmosphere continuum. SPAC in rocky mountain area of Beijing, China. Chinese Journal of Applied Ecology, 7:2171-2178. (in Chinese)
[16]
Ehleringer J R, Dawson T E. 1992. Water uptake by plants: Perspectives from stable isotope composition. Plant Cell and Environment, 15(9):1073-1082.
[17]
Ellsworth P Z, Williams D G. 2007. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant and Soil, 291:93-107.
[18]
Ellsworth P Z, Sternberg L S L. 2015. Seasonal water use by deciduous and evergreen woody species in a scrub community is based on water availability and root distribution. Ecohydrology, 8(4):538-551.
[19]
Herczeg A L, Leaney F W. 2011. Review: Environmental tracers in arid-zone hydrology. Hydrogeology Journal, 19(1):17-29.
[20]
Hirl R T, Schnyder H, Ostler U, et al. 2019. The 18O ecohydrology of a grassland ecosystem-predictions and observations . Hydrology and Earth System Sciences Discussions, 23:2581-2600.
[21]
Horton J L, Hart S C, Kolb T E. 2003. Physiological condition and water source use of Sonoran Desert riparian trees at the Bill Williams River, Arizona, USA. Isotopes in Environmental and Health Studies, 39(1):69-82.
doi: 10.1080/1025601031000096772
pmid: 12812256
[22]
Hu H Y, Li H X, Ni B, et al. 2019. Characteristic of typical vegetation community and water use efficiency of dominant plants in desert steppe of Ningxia. Journal of Zhejiang University (Agriculture & Life Science), 45(4):460-471. (in Chinese)
[23]
Huang L, Zhang Z S. 2015. Stable isotopic analysis on water utilization of two xerophytic shrubs in a revegetated desert area: Tengger Desert, China. Water, 7(3):1030-1045.
[24]
Moreno-Gutiérrez C, Dawson T E, Nicolás E, et al. 2012. Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New Phytologist, 196(2):489-496.
[25]
Nie Y P, Chen H S, Wang K L, et al. 2014. Seasonal variations in leaf δ 13C values: implications for different water-use strategies among species growing on continuous dolomite outcrops in subtropical China . Acta Physiology Plant, 36:2571-2579.
[26]
Oerter E J, Siebert G, Bowling D R, et al. 2019. Soil water vapour isotopes identify missing water source for streamside trees. Ecohydrology 12:e2083, doi: 10.1002/eco.2083.
[27]
Philips D L, Gregg J W. 2003. Source partitioning using stable isotopes: coping with too many sources. Oecologia, 136:261-269.
doi: 10.1007/s00442-003-1218-3
pmid: 12759813
[28]
Querejeta J I, Estrada-Medina H, Allen M F, et al. 2007. Water source partitioning among trees growing on shallow karst soils in a seasonally dry tropical climate. Oecologia, 152:26-36.
doi: 10.1007/s00442-006-0629-3
pmid: 17216213
[29]
Song N P, Yang M X, Wang L, et al. 2014. Monthly variation in soil moisture under Caragana intermedia stands grown in desert steppe. Chinese Journal of Ecology, 33(10):2618-2624. (in Chinese)
[30]
Tanaka-Oda A, Endo I, Ohte N, et al. 2018. A water acquisition strategy may regulate the biomass and distribution of winter forage species in cold Asian rangeland. Ecosphere, 9(12):e02511, doi: 10.1002/ecs2.2511.
[31]
Wang X, Hao Z, Ye J, et al. 2008. Spatial pattern of diversity in an old growth temperate forest in Northeastern China. Acta Oecologica, 33(3):345-354.
[32]
Werner C, Schnyder H, Cuntz M, et al. 2012. Progress and challenges in using stable isotopes to trace plant carbon and water relations across scales. Biogeosciences, 9(8):3083-3111.
[33]
Wittmer M H O M, Auerswald K, Tungalag R, et al. 2008. Carbon isotope discrimination of C3 vegetation in Central Asian grassland as related to long-term and short-term precipitation patterns. Biogeosciences, 5:913-924.
[34]
Wu H W, Li X Y, Li J, et al. 2016. Differential soil moisture pulse uptake by coexisting plants in an alpine Achnatherum splendens grassland community. Environmental Earth Sciences 75:914, doi: 10.1007/s12665-016-5694-2.
[35]
Wu Y, Zhou H, Zheng X J, et al. 2014. Seasonal changes in the water use strategies of three co-occurring desert shrubs. Hydrological Processes, 28(26):6265-6275.
[36]
Yang H, Auerswald K, Bai Y F, et al. 2011. Complementarity in water sources among dominant species in typical steppe ecosystems of Inner Mongolia, China. Plant and Soil, 340:303-313.
[37]
Zhao P P, Shao W S, Jin C Q, et al. 2017. Effects of enclosure on biomass allocation characteristics of Agropyron mongolicum population in desert steppe. Ecology and Environmental Sciences, 26(12):2024-2029. (in Chinese)
[38]
Zhao Y N, Zhou Y R, Wang H M. 2018. Spatial heterogeneity of soil water content under introduced shrub (Caragana korshinskii) in desert grassland of the eastern Ningxia, China. Chinese Journal of Applied Ecology, 29(11):3577-3586. (in Chinese)
doi: 10.13287/j.1001-9332.201811.001
pmid: 30460804
[39]
Zhu L, Zhang H L, Gao X, et al. 2016. Seasonal patterns in water uptake for Medicago sativa grown along an elevation gradient with shallow groundwater table in Yanchi county of Ningxia, Northwest China. Journal of Arid Land, 8(6):921-934.