| Research article |
|


|
|
|
| Competition, spatial pattern, and regeneration of Haloxylon ammodendron and Haloxylon persicum communities in the Gurbantunggut Desert, Northwest China |
LIU Yaxuan1,2, ZENG Yong1,2,*( ), YANG Yuhui1,2, WANG Ning3, LIANG Yuejia1,2 ), YANG Yuhui1,2, WANG Ning3, LIANG Yuejia1,2 |
1Xinjiang Laboratory of Lake Environment and Resources in Arid Zone, Urumqi 830054, China
2College of Geographic Science and Tourism, Xinjiang Normal University, Urumqi 830054, China
3School of Geography and Tourism, Shaanxi Normal University, Xi'an 710119, China |
|
|
|
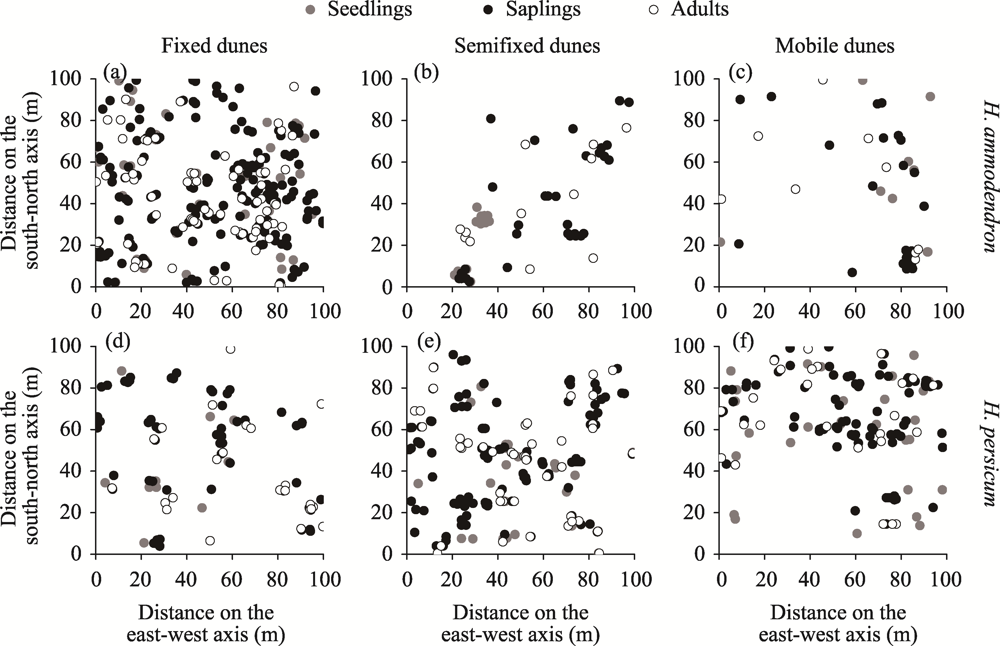
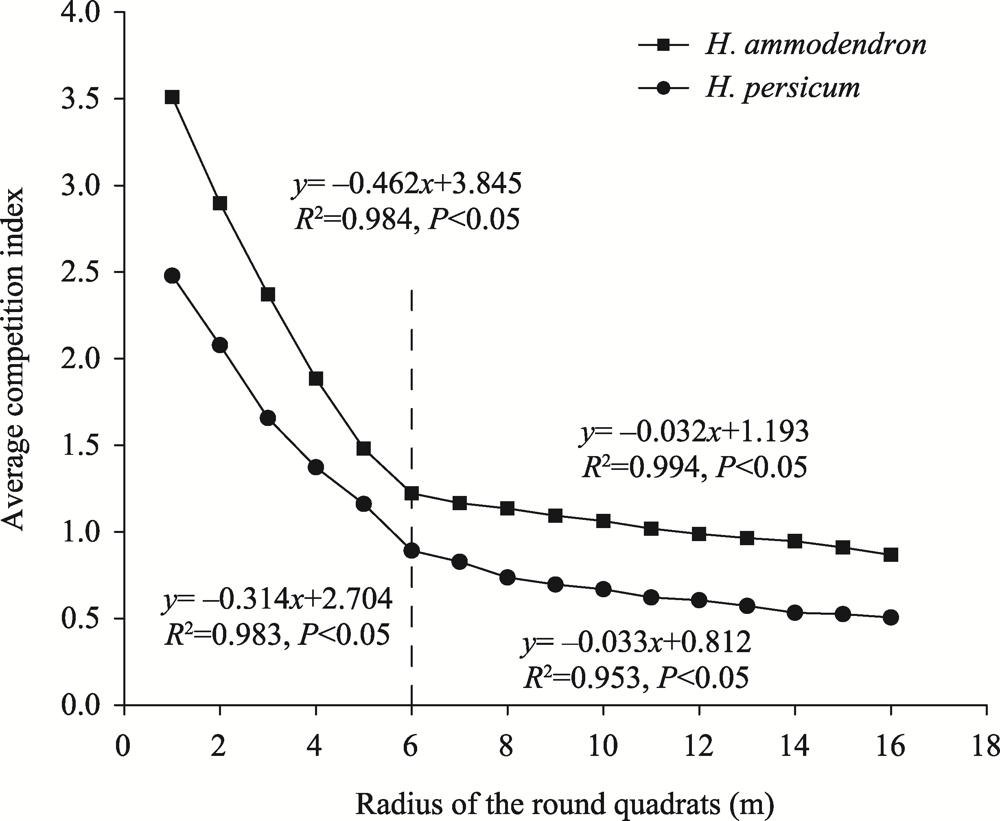
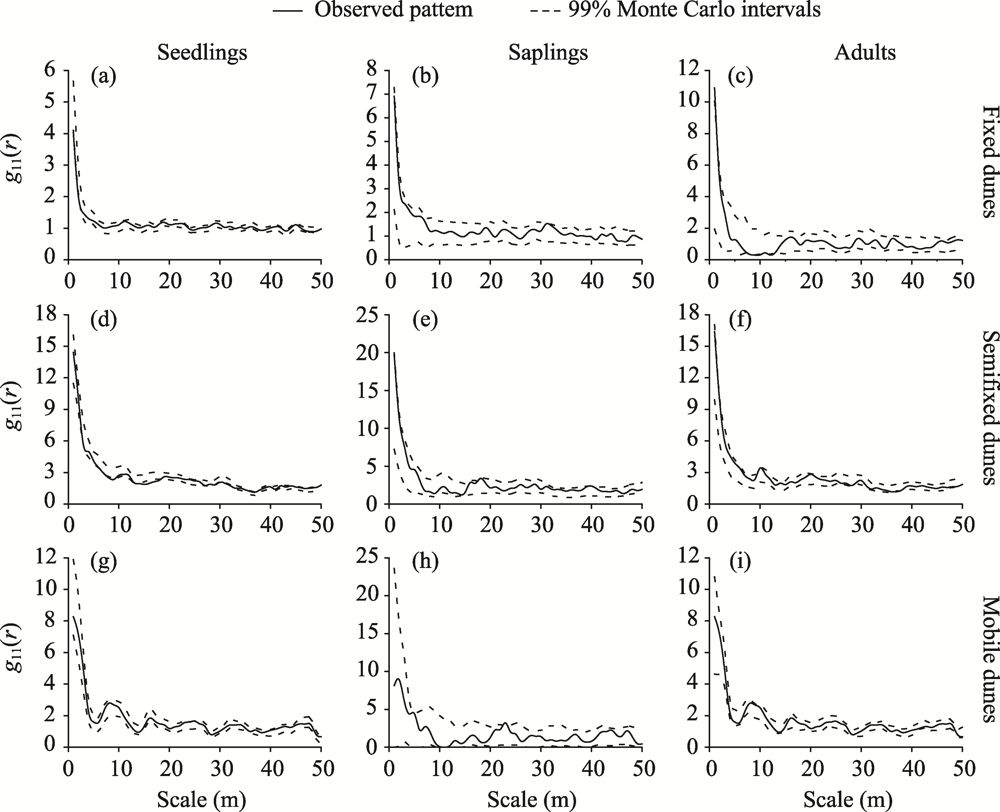
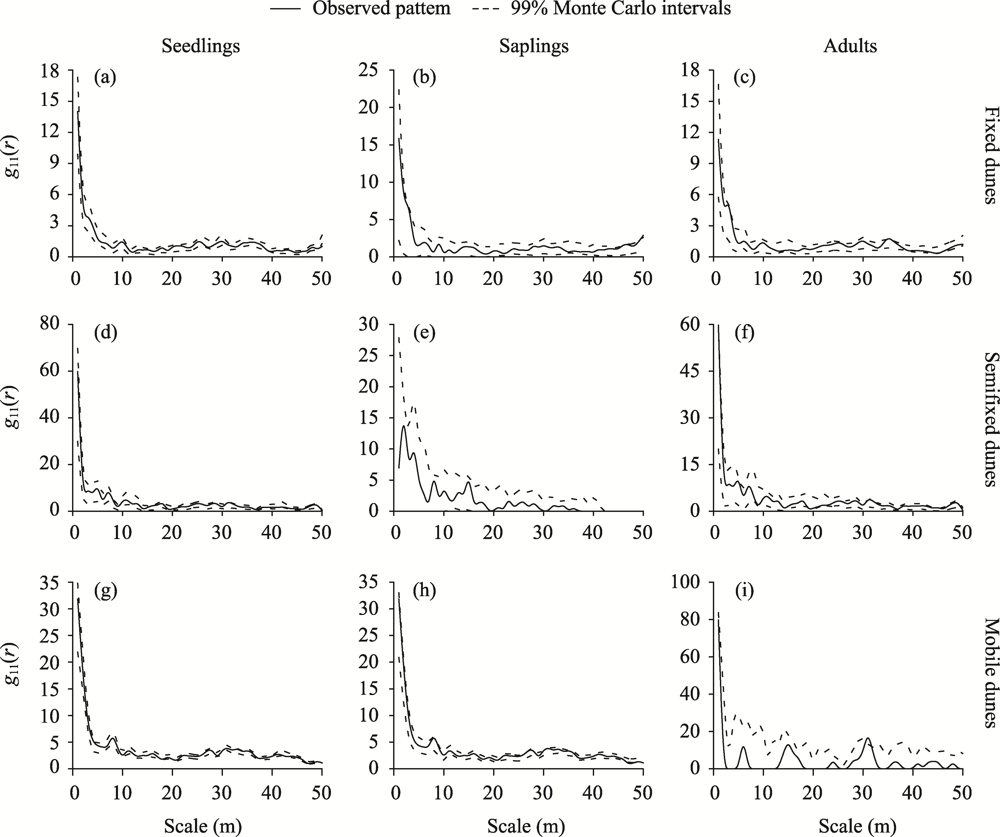
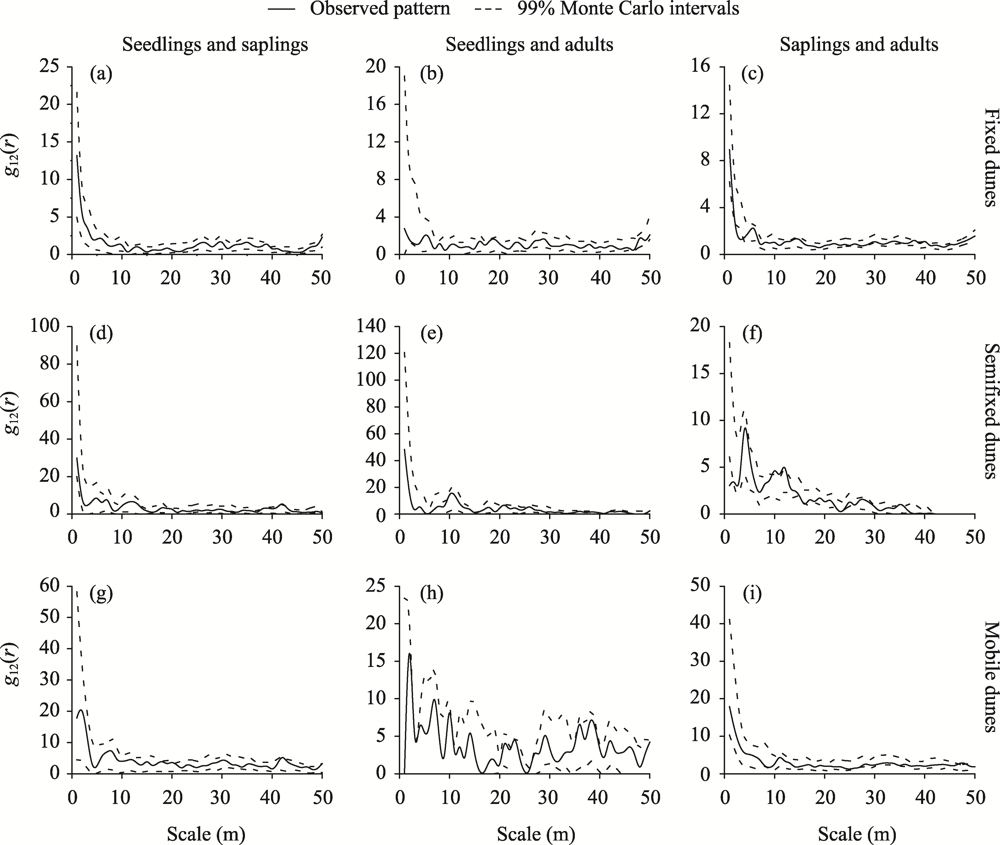
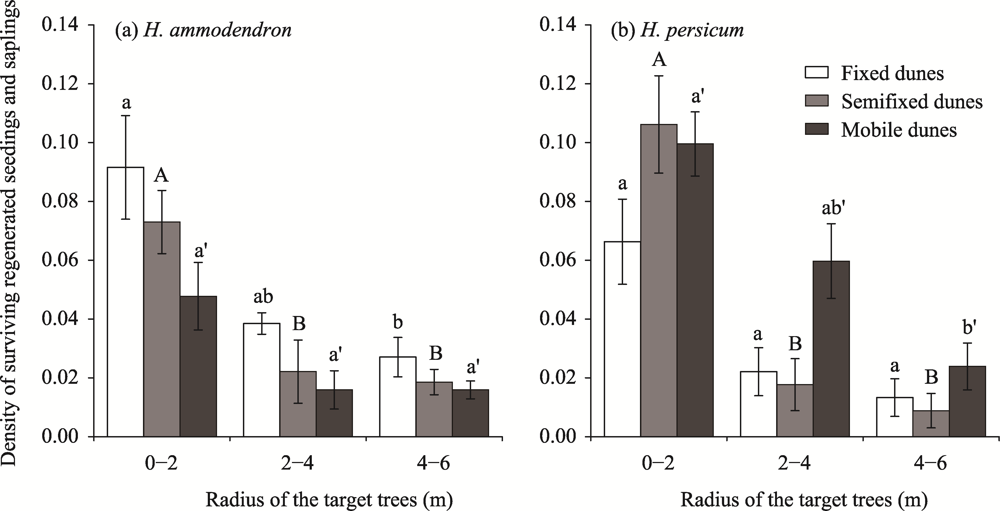
Abstract Competition, spatial pattern, and regeneration are important factors affecting community composition, structure, and dynamics. In this study, we surveyed 300 quadrats from three dunes (i.e., fixed dunes, semifixed dunes, and mobile dunes) in the Gurbantunggut Desert, Northwest China, from late May to early June in 2021. The intraspecific and interspecific competition, spatial pattern, and regeneration of Haloxylon ammodendron and Haloxylon persicum were studied using the Hegyi competition index and point pattern analysis methods. The results showed that the optimal competition distance of the objective tree in the H. ammodendron and H. persicum communities was 6 m. The intraspecific and interspecific competition of H. ammodendron was the greatest in fixed dunes, while the competition intensity of H. persicum in semifixed dunes and mobile dunes was greater than that in fixed dunes. The order of competition intensity of the two populations was seedlings>saplings>adults, and the competition intensity gradually decreased with the increase in plant diameter. The spatial distribution pattern of the three life stages of H. ammodendron and H. persicum was random, and there were no correlations between seedlings and saplings, adults and saplings, and seedlings and adults. The density of regenerated seedlings and saplings of H. ammodendron in the three dunes followed the order of fixed dunes>semifixed dunes>mobile dunes, and that of H. persicum in the three dunes followed the order of mobile dunes>semifixed dunes>fixed dunes. Therefore, when artificially planting H. ammodendron and H. persicum for sand control, the planting interval should be 6 m, and seedlings should be planted next to adults to minimize the competition between plants, which can promote the renewal of H. ammodendron and H. persicum and the stabilization of the ecosystem.
|
|
Received: 21 May 2022
Published: 31 October 2022
|
|
Corresponding Authors:
*ZENG Yong (E-mail: zengyong107@sina.cn)
|
|
|
| [1] |
Adler P B, Smull D, Beard K H, et al. 2018. Competition and coexistence in plant communities: intraspecific competition is stronger than interspecific competition. Ecology Letters, 21(9): 1319-1329.
doi: 10.1111/ele.13098
pmid: 29938882
|
|
|
| [2] |
Ali A, Dai D, Akhtar K, et al. 2019. Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Global Ecology and Conservation, 20: e00775, doi: 10.1016/j.gecco.2019.e00775.
doi: 10.1016/j.gecco.2019.e00775
|
|
|
| [3] |
Alla A Q, Pasho E. 2015. Growth variability and competition influence on tree growth in Abies borisii-regis forests from south-east Albania. In: The 3rd International Conference on Research and Education-"Challenges Toward the Future". Albania.
|
|
|
| [4] |
Ban W Q, Yan C, Yin L K, et al. 2012. Plant species diversity and dominance population niche characteristics at different sites in southern Gurbantunggut Desert. Journal of Desert Research, 32(6): 1632-1638. (in Chinese)
|
|
|
| [5] |
Chang J, Pan C D, Shi R F. 2006. Analysis on dominant species distribution patterns and relation of Ass. Haloxylon persicum and H. ammodendron. Journal of Xinjiang Agricultural University, 29(2): 26-29. (in Chinese)
|
|
|
| [6] |
Chao L, Liu Y Y, Zhang D W. 2017. Diameter class structure, growth dynamic, intraspecific and interspecific competitions of Betula platyphylla on swamp ecotone. Chinese Journal of Ecology, 36(3): 577-583. (in Chinese)
|
|
|
| [7] |
Chazdon R L, Guariguata M R. 2016. Natural regeneration as a tool for large-scale forest restoration in the tropics: prospects and challenges. Biotropica, 48(6): 716-730.
doi: 10.1111/btp.12381
|
|
|
| [8] |
Chen S, Hai X, Shi X W. 2018. The competitive relationship of Pinus massoniana and Quercus aliena mixed forest in Baotianman National Nature Reserve. Journal of Southwest Forestry University (Natural Science), 38(2): 10-15. (in Chinese)
|
|
|
| [9] |
Chen J, Zhang X J, Li Q Y, et al. 2021. Effects of slope direction on the intraspecific and interspecific competition in subalpine color-leaved forests in western Sichuan, China. Journal of Southwest University, 43(2): 53-62. (in Chinese)
|
|
|
| [10] |
Chi S, Wang C J, Li Q J, et al. 2020. Spatial distribution and interspecific associations of regenerating saplings in karst secondary forests. Chinese Journal of Applied Ecology, 31(12): 3989-3996. (in Chinese)
doi: 10.13287/j.1001-9332.202012.006
|
|
|
| [11] |
Craine J M, Dybzinski R. 2013. Mechanisms of plant competition for nutrients, water and light. Functional Ecology, 27(4): 833-840.
doi: 10.1111/1365-2435.12081
|
|
|
| [12] |
Cui Y H. 2015. Spatial distribution patterns and associations of tree species in spruce secondary forest of Guandi Mountain. MSc Thesis. Jinzhong: Shanxi Agricultural University. (in Chinese)
|
|
|
| [13] |
Dai Y, Zheng X J, Li Y, et al. 2013. Stemflow of Haloxylon ammodendron and H. persicum in the Gurbantunggut Desert. Arid Zone Research, 30(5): 867-872. (in Chinese)
|
|
|
| [14] |
D'Amato A W, Orwig D A, Foster D R. 2009. Understory vegetation in old-growth and second-growth Tsuga canadensis forests in western Massachusetts. Forest Ecology and Management, 257(3): 1043-1052.
doi: 10.1016/j.foreco.2008.11.003
|
|
|
| [15] |
Dale M R T. 1999. Spatial pattern analysis in plant ecology. Ecology, 88: 366-370.
|
|
|
| [16] |
Dang J J, Zhao C Z, Dong X, et al. 2014. Response of Kobresia tibetica and Kobresia humilis population spatial pattern to soil moisture. Chinese Journal of Ecology, 33(7): 1734-1740. (in Chinese)
|
|
|
| [17] |
Feng W. 2017. Fine-scale spatial pattern and spatial association of Tamarix chinensis size structure in Qin Wangchuan wetland. MSc Thesis. Lanzhou: Northwest Normal University. (in Chinese)
|
|
|
| [18] |
Gao F Y, Zhao C Z, Zhuo Ma L C. 2014. Spatial distribution and spatial association of Stellera chamaejasme population in the different altitude in degraded alpine grassland. Acta Ecologica Sinica, 34(3): 605-612. (in Chinese)
|
|
|
| [19] |
Gao H J, Gao P S, Wang G M. 2017. Intraspecific and interspecific competition of Machilus thunbergii forest in Zhoushan Islands. Bulletin of Botanical Research, 37(3): 440-446. (in Chinese)
|
|
|
| [20] |
Guo Q S, Guo Z H, Yan H, et al. 2005. Study on potential distribution of Haloxylon plants dominated desert vegetation in China. Acta Ecologica Sinica, 25(4): 848-853. (in Chinese)
|
|
|
| [21] |
He Y, Ding G D, Liang W J, et al. 2011. Study on structural characteristics of shrub community in typical natural secondary forest in Hebei Province. Forest Resources Management, (4): 89-93. (in Chinese)
|
|
|
| [22] |
He A L, Niu S Q, Yang D, et al. 2021. Two PGPR strains from the rhizosphere of Haloxylon ammodendron promoted growth and enhanced drought tolerance of ryegrass. Plant Physiology and Biochemistry, 161: 74-85.
doi: 10.1016/j.plaphy.2021.02.003
|
|
|
| [23] |
Hu X J, Zhang W H, He J F, et al. 2015. Architectural analysis of crown geometry of Quercus variablis BL. natural regenerative seedlings in different habitats. Acta Ecologica Sinica, 35(3): 788-795. (in Chinese)
|
|
|
| [24] |
Hu D, Lv G H, Qie Y D, et al. 2021. Response of morphological characters and photosynthetic characteristics of Haloxylon ammodendron to water and salt stress. Sustainability, 13(1): 388, doi: 10.3390/su13010388.
doi: 10.3390/su13010388
|
|
|
| [25] |
Hu Y C, Zhang X L, Han X Y, et al. 2022. Responses of stand growth, regeneration, and understory species diversity in Quercus mongolica secondary forest to stand density. Chinese Journal of Applied Ecology, 33(3): 727-732. (in Chinese)
|
|
|
| [26] |
Huang X B, Liu W D, Su J R, et al. 2016. Intraspecific and interspecific competition of Pinus yunnanensis natural forest. Forest Research, 29(2): 209-215. (in Chinese)
|
|
|
| [27] |
Ji W J, Hanan N P, Browning D M, et al. 2019. Constraints on shrub cover and shrub-shrub competition in a US southwest desert. Ecosphere, 10(2): e02590, doi: 10.1002/ecs2.2590.
doi: 10.1002/ecs2.2590
|
|
|
| [28] |
Jia Z Q, Lu Q. 2004. Haloxylon Ammodendron (C. A. Mey.) Bunge. Beijing: China Environment Science Press, 63-69. (in Chinese)
|
|
|
| [29] |
Kang D, Deng J, Qin X W, et al. 2017. Effect of competition on spatial patterns of oak forests on the Chinese Loess Plateau. Journal of Arid Land, 9(1): 122-131.
doi: 10.1007/s40333-016-0024-9
|
|
|
| [30] |
Laungani R, Knops J M H. 2009. Species-driven changes in nitrogen cycling can provide a mechanism for plant invasions. Proceedings of the National Academy of Sciences, 106(30): 12400-12405.
|
|
|
| [31] |
Li X L, Wang H, Zheng Z, et al. 2009. Composition, spatial distribution and survival during the dry season of tree seedlings in a tropical forest in Xishuangbanna, SW China. Chinese Journal of Plant Ecology, 33(4): 658-671. (in Chinese)
|
|
|
| [32] |
Li Y K, Zhao C Y, Yang R H. 2016. Intraspecific competition of Haloxylon ammodendron in the southern Margin of Junggar Basin. Journal of Desert Research, 36(2): 335-341. (in Chinese)
|
|
|
| [33] |
Li G L, Zhang D H, Zhang Z S, et al. 2021. Population dynamics of main sand-fixing shrubs in the Gurbantunggut Desert. Journal of Desert Research, 41(2): 129-137. (in Chinese)
|
|
|
| [34] |
Li R, Zhang T, Xie J, et al. 2021. Research progress on the tradeoff between seed regeneration and sprout regeneration and the factors influencing the early process of forest regeneration. Chinese Journal of Ecology, 40(7): 2234-2242. (in Chinese)
|
|
|
| [35] |
Li S, Zhang D H, Zhang Z S, et al. 2022. Spatial distribution pattern and correlation of Artemisia halodendron population in Horqin Sandy Land, China. Biological Resources, 44(1): 63-72. (in Chinese)
|
|
|
| [36] |
Liu H F. 2018. Effects of precipitation changes on seed germination and community diversity of desert plants in Junggar desert. PhD Dissertation. Shihezi: Shihezi University. (in Chinese)
|
|
|
| [37] |
Liu H F, Liu T, Han Z Q, et al. 2018. Germination heterochrony in annual plants of Salsola L.: an effective survival strategy in changing environments. Scientific Reports, 8: 6576, doi: 10.1038/s41598-018-23319-0.
doi: 10.1038/s41598-018-23319-0
|
|
|
| [38] |
Liu W S, Li X, Chen F Y, et al. 2020. Intraspecific and interspecific competition of Quercus mongolica forest. Bulletin of Botanical Research, 40(4): 552-558. (in Chinese)
|
|
|
| [39] |
Long S S, Zeng S Q, Liu F L, et al. 2020. Influence of slope, aspect and competition index on the height-diameter relationship of Cyclobalanopsis glauca trees for improving prediction of height in mixed forests. Silva Fennica, 54(1): 10242, doi: 10.14214/sf.10242.
doi: 10.14214/sf.10242
|
|
|
| [40] |
Lv C Y, Zhang X M, Liu G J, et al. 2012. Population structure and spatial pattern of Haloxylon ammodendron population along northwestern edge of the Junggar Basin. Journal of Desert Research, 32(2): 380-387. (in Chinese)
|
|
|
| [41] |
Morcillo L, Bautista S. 2022. Interacting water, nutrients, and shrub age control steppe grass-on-shrub competition: Implications for restoration. Ecosphere, 13(5): e4093, doi: 10.1002/ecs2.4093.
doi: 10.1002/ecs2.4093
|
|
|
| [42] |
Niu H H, Chen H, Fu Y, et al. 2019. Ecological niche characteristics of desert plants in the eastern Qaidam Basin. Acta Ecologica Sinica, 39(8): 2862-2871. (in Chinese)
|
|
|
| [43] |
Pan L L, Kwon S, Liu Y S, et al. 2019. Tree competition, spatial pattern, and regeneration of a Mongolian pine natural forest in the southern geographical edge. Acta Ecologica Sinica, 39(10): 3687-3699. (in Chinese)
|
|
|
| [44] |
Perry G L W, Miller B P, Enright N J, et al. 2014. Stochastic geometry best explains spatial associations among species pairs and plant functional types in species-rich shrublands. Oikos, 123(1): 99-110.
doi: 10.1111/j.1600-0706.2013.00400.x
|
|
|
| [45] |
Pierce N A, Archer S R, Bestelmeyer B T. 2019. Competition suppresses shrubs during early, but not late, stages of arid grassland-shrubland state transition. Functional Ecology, 33(8): 1480-1490.
doi: 10.1111/1365-2435.13336
|
|
|
| [46] |
Qi K. 2011. Spatial pattern and interspecific associations of main species in a Pinus tabulaeformis forest in Heilihe. MSc Thesis. Beijing: Beijing Forestry University. (in Chinese)
|
|
|
| [47] |
Sardar A K, Hanif M, Asaduzzaman M, et al. 2016. Mathematical analysis of the two species Lotka-Volterra predator-prey inter-specific game theoretic competition model. Advanced Modeling and Optimization, 18(2): 231-242.
|
|
|
| [48] |
Sharma C M, Mishra A K, Krishan R, et al. 2016. Variation in vegetation composition, biomass production, and carbon storage in ridge top forests of high mountains of Garhwal Himalaya. Journal of Sustainable Forestry, 35(2): 119-132.
doi: 10.1080/10549811.2015.1118387
|
|
|
| [49] |
Shen Z Q, Lu J, Hua M, et al. 2016a. Spatial point pattern of Pinus densata forests of Sejila Mountains in Tibet. Journal of Northwest A & F University (Natural Science edition), 44(5): 73-81. (in Chinese)
|
|
|
| [50] |
Shen Z Q, Hua M, Dan Q, et al. 2016b. Spatial pattern analysis and associations of Quercus aquifolioides population at different growth stages in Southeast Tibet, China. Chinese Journal of Applied Ecology, 27(2): 387-394. (in Chinese)
|
|
|
| [51] |
Shi Y F, Zhang Z S, Huang L, et al. 2016. Species composition and population structure of plant communities on semi-fixed dunes of the Gurbantunggut Desert, China. Journal of Applied Ecology, 27(4): 1024-1030. (in Chinese)
|
|
|
| [52] |
Shields J M, Jenkins M A, Saunders M R, et al. 2014. Age distribution and spatial patterning of an invasive shrub in secondary hardwood forests. Forest Science, 60(5): 830-840.
doi: 10.5849/forsci.13-079
|
|
|
| [53] |
Song Y Y, Li Y Y, Zhang W H. 2010. Analysis of spatial pattern and spatial association of Haloxylon ammodendron population in different developmental stages. Acta Ecologica Sinica, 30(16): 4317-4327. (in Chinese)
|
|
|
| [54] |
Song C W, Li C J, Halik Ü et al. 2021. Spatial distribution and structural characteristics for Haloxylon ammodendron plantation on the southwestern edge of the Gurbantunggut Desert. Forests, 12(5): 633, doi: 10.3390/f12050633.
doi: 10.3390/f12050633
|
|
|
| [55] |
Sun Y X, Yang J Y, Qiao Y Q, et al. 2020. Spatial distribution pattern of Reaumuria songarica population in Alashan Plateau. Journal of Desert Research, 40(1): 105-115.
|
|
|
| [56] |
Tian Q L, Liu T. 2020. Relationship between the distribution characteristics of fine roots of Haloxylon persicum and soil moisture under extreme drought conditions. Journal of Shihezi University (Natural Science), 38(1): 75-82. (in Chinese)
|
|
|
| [57] |
Tirado R, Bråthen K A, Pugnaire F I. 2015. Mutual positive effects between shrubs in an arid ecosystem. Scientific Reports, 5: 14710, doi: 10.1038/srep14710.
doi: 10.1038/srep14710
pmid: 26419958
|
|
|
| [58] |
Tobe K, Li X, Omasa K. 2000. Effects of sodium chloride on seed germination and growth of two Chinese desert shrubs, Haloxylon ammodendron and H. persicum (Chenopodiaceae). Australian Journal of Botany, 48(4): 455-460.
doi: 10.1071/BT99013
|
|
|
| [59] |
Wang L, Sun Q W, Hao C Y, et al. 2010. Point pattern analysis of different age-class Taxus chinensis var. mairei individuals in mountainous area of southern Anhui province. Chinese Journal of Applied Ecology, 21(2): 272-278. (in Chinese)
pmid: 20461993
|
|
|
| [60] |
Wang H J, Chang S L, Zhang Y T, et al. 2016. Density-dependent effects in Picea schrenkiana forests in Tianshan Mountains. Biodiversity Science, 24(3): 252-261. (in Chinese)
doi: 10.17520/biods.2015282
|
|
|
| [61] |
Wang X J. 2017. Study on relationship of sand-fixating vegetation pattern and hydrological process in Gurbantunggut Desert. PhD Dissertation. Urumqi: Xinjiang Agricultural University. (in Chinese)
|
|
|
| [62] |
Wang X M, Kang X, Hou C Y, et al. 2017. Influence factors of Ilex chinensis seedling regeneration in the mountainous region of southern Jiangsu Province. Journal of Nanjing Forestry University (Natural Sciences Edition), 60(4): 197-201. (in Chinese)
|
|
|
| [63] |
Wang D W. 2019. The influence of large trees on spatial distribution of adjacent trees in broadleaved Korean pine forests in Xiaoxing'an Mountains. MSc Thesis. Harbin: Northeast Forestry University. (in Chinese)
|
|
|
| [64] |
Wang Q, Pan P, Ouyang X Z, et al. 2021. Intraspecific and interspecific competition intensity in mixed plantation with different proportion of Pinus massoniana and Schima superba. Chinese Journal of Ecology, 40(1): 49-57. (in Chinese)
|
|
|
| [65] |
Wei Y, Yin L K, Yan C. 2005. Study on the flowering and pollination characteristics of Haloxylon persicum. Arid Zone Research, 22(1): 85-89. (in Chinese)
|
|
|
| [66] |
Wiegand T, A,. Moloney K. 2004. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 104(2): 209-229.
doi: 10.1111/j.0030-1299.2004.12497.x
|
|
|
| [67] |
Wu P, Peng X Q, Yang S R, et al. 2019. Spatial distribution patterns and correlation of Tamarix chinensis population in coastal wetlands of Shandong, China. Chinese Journal of Plant Ecology, 43(9): 817-824. (in Chinese)
doi: 10.17521/cjpe.2018.0186
|
|
|
| [68] |
Xia J B, Zhao X M, Liu J H, et al. 2016. Environmental factors influencing the distribution of Tamarix chinensis Lour in the Laizhou Bay wetland of the Yellow River Delta. Acta Ecologica Sinica, 36(15): 4801-4808. (in Chinese)
|
|
|
| [69] |
Xiang X Y, Wu G L, Duan R Y, et al. 2015. Intraspecific and interspecific competition of Pinus dabeshanesis. Acta Ecologica Sinica, 35(2): 389-395. (in Chinese)
|
|
|
| [70] |
Xu G Q, Yu D D, Li Y. 2017. Patterns of biomass allocation in Haloxylon persicum woodlands and their understory herbaceous layer along a groundwater depth gradient. Forest Ecology and Management, 395: 37-47.
doi: 10.1016/j.foreco.2017.03.037
|
|
|
| [71] |
Xu H, Liu Y H. 2018. Relationship between diameter class structure and intraspecific and interspecific competitions of precious and endangering plant Acer catalpifolium. Acta Botanica Boreali-Occidentalia Sinica, 38(6): 1160-1170. (in Chinese)
|
|
|
| [72] |
Xu J W, Shi F X, Zhang C H, et al. 2020. Difference in intra- and inter-specific competition of two endangered plant species (Toona ciliate var. pubescens and Taxus chinensis var. mairei) in the middle subtropical zone of China. The Journal of Applied Ecology, 31(1): 1-8. (in Chinese)
doi: 10.2307/2404594
|
|
|
| [73] |
Yan H, Sun F F, Ma S M, et al. 2021. Population structure and spatial distribution pattern of Haloxylon ammodendron and H. persicum. Southwest China Journal of Agricultural Sciences, 34(8): 1781-1787. (in Chinese)
|
|
|
| [74] |
Yang Y F, Zhou H F, Xu L G. 2011. Dynamic variations of soil moisture in Haloxylon ammodendron root zone in Gurbantunggut Desert. Chinese Journal of Applied Ecology, 22(7): 1711-1716. (in Chinese)
pmid: 22007445
|
|
|
| [75] |
Yang S Q, Yi L T, Ye N N, et al. 2022. Spatial pattern dynamics of Cyclobalanopsis myrsinifolia in mixed broad-leaved forests on Tianmu Mountain, eastern China, 1996-2012. Silva Fennica, 56(1): 10520, doi: 10.14214/sf.10520.
doi: 10.14214/sf.10520
|
|
|
| [76] |
Zeng Y, Liu T, Zhou X B, et al. 2016. Effects of climate change on plant composition and diversity in the Gurbantunggut Desert of northwestern China. Ecological Research, 31(3): 427-439.
doi: 10.1007/s11284-016-1352-0
|
|
|
| [77] |
Zeng Y, Zhao C Y, Li C J, et al. 2019. Spatial distribution pattern and correlation of Populus euphratica communities in different habitats along the Tarim River. Chinese Journal of Ecology, 38(11): 3273-3282. (in Chinese)
|
|
|
| [78] |
Zhang J B, Wang B F, Hao Y G, et al. 2010. Geographical distribution, suitable environment and provenance variation of Haloxylon ammdendron (CAM) Bge in China. Journal of Arid Land Resources and Environment, 24(5): 166-171. (in Chinese)
|
|
|
| [79] |
Zhang Z, Liu P, Ding Y, et al. 2010. Distribution patterns of Picea schrenkiana var. tianshanica populations at different developmental stages in the western Tianshan Mountain, northwestern China. Journal of Beijing Forestry University, 32(3): 75-79. (in Chinese)
|
|
|
| [80] |
Zhang X B, Shangguan Z P, Wang J C, et al. 2014. The regeneration characteristics and affecting factors of Pinus tabulaeformis artificial forests of Ziwu Mountains, China. Mountain Research, 32(5): 561-567. (in Chinese)
|
|
|
| [81] |
Zhang S Z, Li M, Zhang S B, et al. 2015. Factors affecting natural regeneration of Larix principis-rupprechtii plantations in Saihanba of Hebei, China. Acta Ecologica Sinica, 35(16): 5403-5411. (in Chinese)
|
|
|
| [82] |
Zhang G F, Yao R Q, Jiang Y, et al. 2016. Intraspecific and interspecific competition intensity of Parrotia subaequalis in different habitats from Wanfoshan Nature Reserve, Anhui Province. Chinese Journal of Ecology, 35(7): 1744-1750. (in Chinese)
|
|
|
| [83] |
Zhang P J, Qing H, Zhao L Q, et al. 2021. Spatial pattern and association of shrub species in gravel hilly and rocky low mountain desert dominated by relict Helianthemum songaricum in China. Global Ecology and Conservation, 32: e01914, doi: 10.1016/j.gecco.2021.e01914.
doi: 10.1016/j.gecco.2021.e01914
|
|
|
| [84] |
Zhang L, Gao Y, Li J R, et al. 2022. Effects of grazing disturbance of spatial distribution pattern and interspecies relationship of two desert shrubs. Journal of Forestry Research, 33(2): 507-518.
doi: 10.1007/s11676-021-01353-5
|
|
|
| [85] |
Zhang Y L, Lu Y X, Yin B F, et al. 2022. Effects of simulated rainfall on soil nutrient contents and enzyme activities in the Gurbantunggut Desert, China. Acta Ecoclogica Sinica, 42(5): 1739-1749. (in Chinese)
|
|
|
| [86] |
Zheng Y, Zhao W Z, Zhang G F. 2017. Spatial analysis of competition in Haloxylon ammodendron community based on the V-Hegyi index in an oasis-desert ecotone. Journal of Desert Research, 37(6): 1127-1134. (in Chinese)
|
|
|
| [87] |
Zhu W R, Li W H, Shi P L, et al. 2021. Intensified interspecific competition for water after afforestation with Robinia pseudoacacia into a native shrubland in the Taihang Mountains, northern China. Sustainability, 13: 807, doi: 10.3390/su13020807.
doi: 10.3390/su13020807
|
|
|
| [88] |
Zhu Z X. 2021. Influencing factors and quantitative model of regeneration for natural Larix gmelinii secondary forest in central Xing'an range. MSc Thesis. Harbin: Northeastern Forestry University. (in Chinese)
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|