| Research article |
|


|
|
|
| Effect of drought and elevated temperature on the physiological and biochemical properties of C3 and C4 halophytes in Amaranthaceae |
Zulfira RAKHMANKULOVA1, Elena SHUYSKAYA1, Maria PROKOFIEVA1, Kristina TODERICH2,3,*( ), Luizat SAIDOVA1, ZHANG Yuanming4 ), Luizat SAIDOVA1, ZHANG Yuanming4 |
1Timiryazev Institute of Plant Physiology, Russian Academy of Sciences, Moscow 127276, Russia
2International Platform for Drylands Research and Education, Tottori 680-0000, Japan
3Institute of Agrobiotechnologies and Food Security, Samarkand State University, Samarkand 140104, Uzbekistan
4Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China |
|
|
|
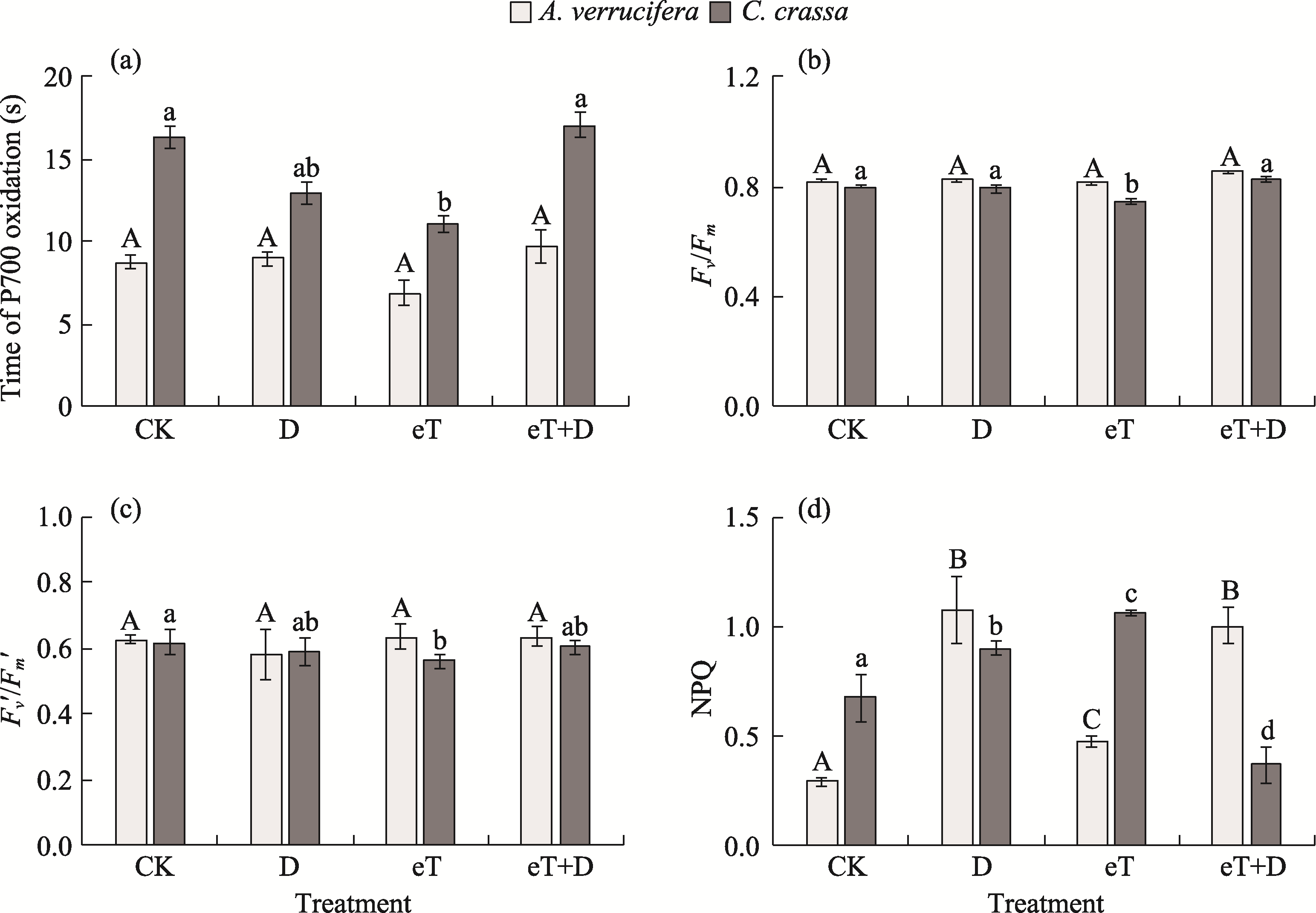
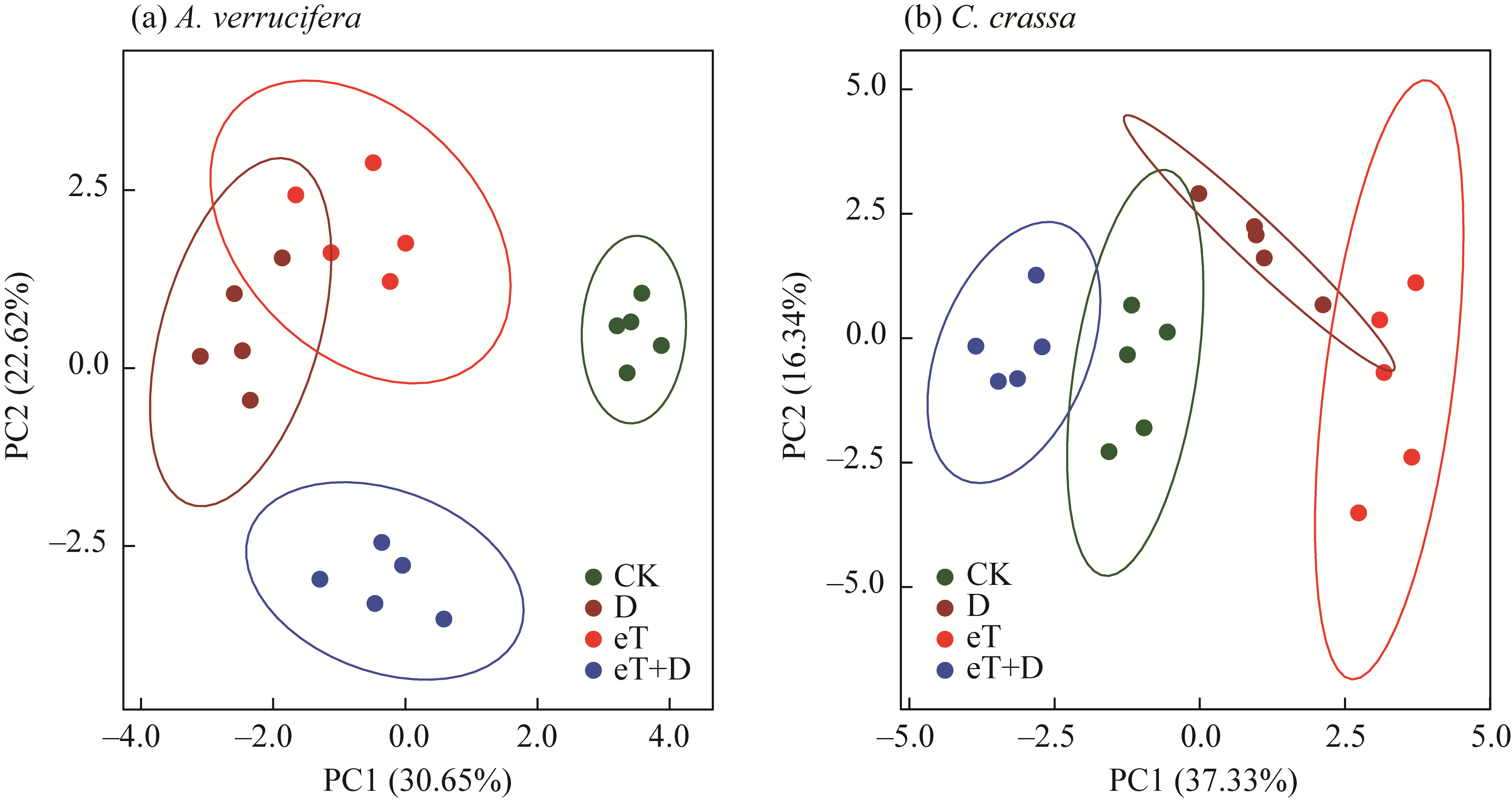
Abstract Rising temperatures and increased droughts caused by climate change significantly reduce crop yields. Halophytes with different photosynthetic metabolism types have specific mechanisms for resistance to climatic factors. This study analyzed the morphophysiological, biochemical, and molecular-genetic mechanisms of tolerance and adaptation in halophytes, promising candidates for the restoration of salt affected lands in arid and semi-arid areas. Experiments under drought (D) and elevated temperature (eT), as well as their combined action (eT+D), were performed on Atriplex verrucifera M. Bied. (C3 plant) and Climacoptera crassa (M. Bieb.) Botsch. (C4-NAD-ME plant) with different types of photosynthesis. The activity of photosystem I (PSI) and the efficiency of photosystem II (PSII) were measured, along with the expression of genes involved in the light (psaA, psaB, psbA, CAB, Fd1, PGR5, and ndhH) and dark (rbcL, Ppc2, and PPDK) reactions of photosynthesis. The content of key carboxylating enzymes ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) and phosphoenolpyruvate carboxylase (PEPC), as well as the photorespiration enzyme glycine decarboxylase (GDC), were assessed. Plant growth and water-salt balance parameters, and activity of enzymes in the malate dehydrogenase (MDH) system nicotinamide adenine dinucleotide (phosphate) (NAD(P))-MDH and NAD(P)-malic enzyme (ME) were also examined. A multivariate analysis of the experimental results revealed that A. verrucifera and C. crassa were both resistant to the effects of these climatic stressors. The tolerance mechanisms of both species were significantly influenced by a high level of photosynthetic plasticity. Nevertheless, differences were observed in the protective mechanisms underlying tolerance. In the C3 species, dissipative processes associated with non-photochemical quenching (NPQ) of PSII and MDH system enzymes (malate valves) were activated, particularly under osmotic stress. The negative effects in the C3 plants were caused by the combined action of eT+D, which was compensated by an increased expression of rbcL, psaA, CAB, and especially PGR5, i.e., genes encoding Rubisco large subunit and PSI components: apoproteins A, chlorophyll a/b-associated protein (CAB) of light-harvesting complex, and proton gradient regulation 5 (PGR5) protein of the main pathway of cyclic electron transport (CET) around PSI. In C4 species, the protective MDH complex was expressed to a lesser extent, but activation of the C4 carbon-concentrating mechanism (CCM) and upregulation of PGR5 expression were observed, particularly under the individual action of the factors. Under the combined stress of eT+D, C. crassa exhibited a synergistic effect, where the increase in NPQ level and NAD-ME activity, as well as decrease in NADP-ME activity was less pronounced compared with the effect of singular factors. Comparative physiological, biochemical, and molecular analyses of how C3 and C4 species response to individual and combined climatic factors provide new insights into sustainable plant adaptation strategies in the face of global climate change. Considering the high nutritional value of these two fodder species, a technological approach could be developed to improve the productivity of salt affected lands.
|
|
Received: 22 August 2025
Published: 31 January 2026
|
|
Corresponding Authors:
*Kristina TODERICH (E-mail: ktoderich@bio.mie-u.ac.jp; ktoderich@yahoo.com)
|
|
|
| [1] |
Alfonso S U, Brüggemann W. 2012. Photosynthetic responses of C3 and three C4 species of the genus Panicum (s.l.) with different metabolic subtypes to drought stress. Photosynthesis Research, 112: 175-191.
doi: 10.1007/s11120-012-9763-4
|
|
|
| [2] |
Alric J, Johnson X. 2017. Alternative electron transport pathways in photosynthesis: a confluence of regulation. Current Opinion in Plant Biology, 37: 78-86.
doi: S1369-5266(16)30188-1
pmid: 28426976
|
|
|
| [3] |
Ashraf M, Harris P J C. 2013. Photosynthesis under stressful environments: An overview. Photosynthetica, 51(2): 163-190.
doi: 10.1007/s11099-013-0021-6
|
|
|
| [4] |
Babayev H, Mehvaliyeva U, Aliyeva M, et al. 2014. The study of NAD-malic enzyme in Amaranthus cruentus L. under drought. Plant Physiology and Biochemistry, 81: 84-89.
doi: 10.1016/j.plaphy.2013.12.022
pmid: 24444721
|
|
|
| [5] |
Bates L S, Waldren R P, Teare I D. 1973. Rapid determination of free proline for water stress studies. Plant Soil, 39: 205-207.
doi: 10.1007/BF00018060
|
|
|
| [6] |
Bianconi M E, Sotelo G, Curran E V, et al. 2022. Upregulation of C4 characteristics does not consistently improve photosynthetic performance in intraspecific hybrids of a grass. Plant, Cell & Environment, 45(5): 1398-1411.
doi: 10.1111/pce.v45.5
|
|
|
| [7] |
Brestic M, Zivcak M. 2013. PSII fluorescence techniques for measurement of drought and high temperature stress signal in crop plants: protocols and applications.In: RoutG R, DasBandhu A. Molecular Stress Physiology of Plants. New Delhi: Springer, 87-131.
|
|
|
| [8] |
Brignone N F, Pozner R, Denham S S. 2022. Macroevolutionary trends and diversification dynamics in Atripliceae (Amaranthaceae s.l., Chenopodioideae): a first approach. Annals of Botany, 130(2): 199-214.
doi: 10.1093/aob/mcac085
pmid: 35737947
|
|
|
| [9] |
Cacefo V, Ribas A F, Zilliani R R, et al. 2019. Decarboxylation mechanisms of C4 photosynthesis in Saccharum spp.: increased PEPCK activity under water-limiting conditions. BMC Plant Biology, 19(1): 144, doi: 10.1186/s12870-019-1745-7.
|
|
|
| [10] |
Calone R, Cellini A, Manfrini L, et al. 2021. The C4 Atriplex halimus vs. the C3 Atriplex hortensis: Similarities and differences in the salinity stress response. Agronomy, 11(10): 1967, doi: 10.3390/agronomy11101967.
|
|
|
| [11] |
Carmo-Silva A E, Bernardes da Silva A, Keys A J, et al. 2008. The activities of PEP carboxylase and the C4 acid decarboxylases are little changed by drought stress in three C4 grasses of different subtypes. Photosynthesis Research, 97(3): 223-233.
doi: 10.1007/s11120-008-9329-7
pmid: 18629606
|
|
|
| [12] |
Carmo-Silva A E, Salvucci M E. 2012. The temperature response of CO2 assimilation, photochemical activities and Rubisco activation in Camelina sativa, a potential bioenergy crop with limited capacity for acclimation to heat stress. Planta, 236(5): 1433-1445.
doi: 10.1007/s00425-012-1691-1
pmid: 22733425
|
|
|
| [13] |
Chadee A, Alber N A, Dahal K, et al. 2021. The complementary roles of chloroplast cyclic electron transport and mitochondrial alternative oxidase to ensure photosynthetic performance. Frontiers in Plant Science, 12: 748204, doi: 10.3389/fpls.2021.748204.
|
|
|
| [14] |
Chen Q Q, Wang B P, Ding H Y, et al. 2019. Review: The role of NADP-malic enzyme in plants under stress. Plant Science, 281: 206-212.
doi: S0168-9452(18)31029-X
pmid: 30824053
|
|
|
| [15] |
Chen S M, Peng W M H, Ansah E O, et al. 2022. Encoded C4 homologue enzymes genes function under abiotic stresses in C3 plant. Plant Signaling & Behavior, 17(1): 2115634, doi: 10.1080/15592324.2022.2115634.
|
|
|
| [16] |
Degen G E, Orr D J, Carmo-Silva E. 2021. Heat-induced changes in the abundance of wheat Rubisco activase isoforms. New Phytologist, 229: 1298-1311.
doi: 10.1111/nph.v229.3
|
|
|
| [17] |
Demircan N, Sonmez M C, Akyol T Y, et al. 2024. Alternative electron sinks in chloroplasts and mitochondria of halophytes as a safety valve for controlling ROS production during salinity. Physiologia Plantarum, 176(3): e14397, doi: 10.1111/ppl.14397.
|
|
|
| [18] |
Dietz K J. 2021. Synergism and antagonism in plant acclimation to abiotic stress combinations. Turkish Journal of Botany, 45(1): 587-600.
doi: 10.3906/bot-2108-45
|
|
|
| [19] |
Doubnerová Hýsková V, Miedzińska L, Dobrá J, et al. 2014. Phosphoenolpyruvate carboxylase, NADP-malic enzyme, and pyruvate, phosphate dikinase are involved in the acclimation of Nicotiana tabacum L. to drought stress. Journal of Plant Physiology, 171(5): 19-25.
doi: 10.1016/j.jplph.2013.10.017
pmid: 24484954
|
|
|
| [20] |
Edwards G E, Voznesenskaya E V. 2010. Chapter 4 C4 photosynthesis: kranz forms and single-cell C4 in terrestrial plants. In: Raghavendra A S, Sage R F. C4 Photosynthesis and Related CO2 Concentrating Mechanisms. Dordrecht: Springer, 29-61.
|
|
|
| [21] |
Elsässer M, Feitosa-Araujo E, Lichtenauer S, et al. 2020. Photosynthetic activity triggers pH and NAD redox signatures across different plant cell compartments. [2025-04-28].https://www.biorxiv.org/content/10.1101/2020.10.31.363051v1.full.pdf.
|
|
|
| [22] |
Eprintsev A T, Fedorina O S. 2007. Functioning of malate dehydrogenase system in mesophyll and bundle sheath cells of maize leaves under salt stress conditions. Russian Journal of Plant Physiology, 54(6): 728-735.
doi: 10.1134/S1021443707060027
|
|
|
| [23] |
Fan Y Z, Asao S, Furbank R T, et al. 2022. The crucial roles of mitochondria in supporting C4 photosynthesis. New Phytologist, 233(3): 1083-1096.
doi: 10.1111/nph.v233.3
|
|
|
| [24] |
Flowers T J, Colmer T D. 2008. Salinity tolerance in halophytes. New Phytologist, 179(4): 945-963.
doi: 10.1111/j.1469-8137.2008.02531.x
pmid: 18565144
|
|
|
| [25] |
Flowers T J, Colmer T D. 2015. Plant salt tolerance: adaptations in halophytes. Annals of Botany, 115(3): 327-331.
pmid: 25844430
|
|
|
| [26] |
Furbank R T. 2017. Walking the C4 pathway: past, present, and future. Journal of Experimental Botany, 67(2): 4057-4066.
doi: 10.1093/jxb/erw161
|
|
|
| [27] |
Ghannoum O, von Caemmerer S, Conroy J P. 2002. The effect of drought on plant water use efficiency of nine NAD-ME and nine NADP-ME C4 Australian grasses. Functional Plant Biology, 29(11): 1337-1348.
doi: 10.1071/FP02056
pmid: 32688732
|
|
|
| [28] |
Ghannoum O, Evans J R, Chow W S, et al. 2005. Faster Rubisco is the key to superior nitrogen-use efficiency in NADP-malic enzyme relative to NAD-malic enzyme C4 grasses. Plant Physiology, 137(2): 638-650.
doi: 10.1104/pp.104.054759
pmid: 15665246
|
|
|
| [29] |
Ghannoum O. 2009. C4 photosynthesis and water stress. Annals of Botany, 103(4): 635-644.
doi: 10.1093/aob/mcn093
pmid: 18552367
|
|
|
| [30] |
Hamed K B, Ellouzi H, Talbi O Z, et al. 2013. Physiological response of halophytes to multiple stresses. Functional Plant Biology, 40(9): 883-896.
doi: 10.1071/FP13074
pmid: 32481158
|
|
|
| [31] |
Hasanuzzaman M, Raihan M R H, Masud A A C, et al. 2021. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. International Journal of Molecular Sciences, 22(17): 9326, doi: 10.3390/ijms22179326.
|
|
|
| [32] |
Hatch M D. 1971. The C4 pathway of photosynthesis. Evidence for an intermediate pool of carbon dioxide and the identity of the donor C4-dicarboxylic acid. Biochemical Journal, 125(2): 425-432.
pmid: 5144745
|
|
|
| [33] |
Hatch M D, Tsuzuki M, Edwards G E. 1982. Determination of NAD malic enzymes in leaves of C(4) plants: effects of malate dehydrogenase and other factors. Plant Physiology, 69(2): 483-491.
doi: 10.1104/pp.69.2.483
pmid: 16662234
|
|
|
| [34] |
Hatch M D. 1987. C4 photosynthesis: a unique elend of modified biochemistry, anatomy and ultrastructure. Biochimica et Biophysica Acta (BBA) - Reviews on Bioenergetics, 895(2): 81-106.
doi: 10.1016/S0304-4173(87)80009-5
|
|
|
| [35] |
Heath R L, Packer L. 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Archives of Biochemistry and Biophysics, 125(1): 180-198.
|
|
|
| [36] |
Ishikawa N, Takabayashi A, Sato F, et al. 2016. Accumulation of the components of cyclic electron flow around photosystem I in C4 plants, with respect to the requirements for ATP. Photosynthesis Research, 129(3): 261-277.
doi: 10.1007/s11120-016-0251-0
pmid: 27017612
|
|
|
| [37] |
Jacoby R P, Taylor N L, Millar A H. 2011. The role of mitochondrial respiration in salinity tolerance. Trends in Plant Science, 16(11): 614-623.
doi: 10.1016/j.tplants.2011.08.002
pmid: 21903446
|
|
|
| [38] |
Jithesh M N, Prashanth S R, Sivaprakash K R, et al. 2006. Antioxidative response mechanisms in halophytes: their role in stress defence. Journal of Genetics, 85(3): 237-254.
pmid: 17406103
|
|
|
| [39] |
Koteyeva N K, Voznesenskaya E V, Edwards G E. 2015. An assessment of the capacity for phosphoenolpyruvate carboxykinase to contribute to C4 photosynthesis. Plant Science, 235: 70-80.
doi: 10.1016/j.plantsci.2015.03.004
pmid: 25900567
|
|
|
| [40] |
Krämer M, Kunz H-H. 2021. Indirect export of reducing equivalents from the chloroplast to resupply NADP for C3 photosynthesis—growing importance for stromal NAD(H)? Frontiers in Plant Science, 12: 719003, doi: 10.3389/fpls.2021.719003.
|
|
|
| [41] |
Krieger-Liszkay A, Shimakawa G. 2022. Regulation of the generation of reactive oxygen species during photosynthetic electron transport. Biochemical Society Transactions, 50(2): 1025-1034.
doi: 10.1042/BST20211246
pmid: 35437580
|
|
|
| [42] |
Lawlor D W. 1995. Effects of water deficit on photosynthesis. In: SmirnoffM. Environment and Plant Metabolism:Flexibility and Acclimation. Oxford: Bios Scientific Publishers, 129-160.
|
|
|
| [43] |
Le X Y H, Millar A H. 2023. The diversity of substrates for plant respiration and how to optimize their use. Plant Physiology, 191(4): 2133-2149.
doi: 10.1093/plphys/kiac599
|
|
|
| [44] |
Lee S C, Luan S. 2012. ABA signal transduction at the crossroad of biotic and abiotic stress responses. Plant, Cell & Environment, 35(1): 53-60.
doi: 10.1111/pce.2012.35.issue-1
|
|
|
| [45] |
Lin H C, Arrivault S, Coe R A, et al. 2020. A partial C4 photosynthetic biochemical pathway in rice. Frontiers in Plant Science, 11: 564463, doi: 10.3389/fpls.2020.564463.
|
|
|
| [46] |
Maier A, Zell M B, Maurino V G. 2011. Malate decarboxylases: evolution and roles of NAD(P)-ME isoforms in species performing C(4) and C(3) photosynthesis. Journal of Experimental Botany, 62(9): 3061-3069.
doi: 10.1093/jxb/err024
pmid: 21459769
|
|
|
| [47] |
Martins-Noguerol R, Rico-Jiménez D, Matías L, et al. 2024. Effects of drought and increased temperature on phytochemical traits of the edible halophyte Crithmum maritimum: Perspectives for future climatic scenarios. Environmental and Experimental Botany, 226: 105924, doi: 10.1016/j.envexpbot.2024.105924.
|
|
|
| [48] |
Munekage Y, Hashimoto M, Miyake C, et al. 2004. Cyclic electron flow around photosystem I is essential for photosynthesis. Nature, 429: 579-582.
doi: 10.1038/nature02598
|
|
|
| [49] |
Nakamura N, Iwano M, Havaux M, et al. 2013. Promotion of cyclic electron transport around photosystem I during the evolution of NADP-malic enzyme‐type C4 photosynthesis in the genus Flaveria. New Phytologist, 199(3): 832-842.
doi: 10.1111/nph.12296
pmid: 23627567
|
|
|
| [50] |
Opoku E, Sahu P P, Findurová H, et al. 2024. Differential physiological and production responses of C3 and C4 crops to climate factor interactions. Frontiers in Plant Science, 15: 1345462, doi: 10.3389/fpls.2024.1345462.
|
|
|
| [51] |
Panda S K, Gupta D, Patel M, et al. 2024. Functionality of reactive oxygen species (ROS) in plants: Toxicity and control in Poaceae crops exposed to abiotic stress. Plants, 13(15): 2071, doi: 10.3390/plants13152071.
|
|
|
| [52] |
Perdomo J A, Capó-Bauçà S, Carmo-Silva E, et al. 2017. Rubisco and Rubisco activase play an important role in the biochemical limitations of photosynthesis in rice, wheat, and maize under high temperature and water deficit. Frontiers in Plant Science, 8: 490, doi: 10.3389/fpls.2017.00490.
pmid: 28450871
|
|
|
| [53] |
Pyankov V I, Kuzmin A N, Demidov E D, et al. 1992. Diversity of biochemical pathways of CO2 fixation in plants of the families Poaceae and Chenopodiaceae from arid zone of Central Asia. Soviet Plant Physiology, 39(4): 411-420.
|
|
|
| [54] |
Qiu N, Lu C. 2003. Enhanced tolerance of photosynthesis against high temperature damage in salt-adapted halophyte Atriplex centralasiatica plants. Plant, Cell & Environment, 26(7): 1137-1145.
doi: 10.1046/j.1365-3040.2003.01038.x
|
|
|
| [55] |
Rakhmankulova Z F, Shuyskaya E V, Shcherbakov A V, et al. 2015. Content of proline and flavonoids in the shoots of halophytes inhabiting the South Urals. Russian Journal of Plant Physiology, 62(1): 71-79.
doi: 10.1134/S1021443715010112
|
|
|
| [56] |
Rao X L, Dixon R A. 2016. The differences between NAD-ME and NADP-ME subtypes of C4 photosynthesis: more than decarboxylating enzymes. Frontiers in Plant Science, 7: 1525, doi: 10.3389/fpls.2016.01525.
|
|
|
| [57] |
Rao X L, Dixon R A. 2019. Corrigendum: The differences between NAD-ME and NADP-ME subtypes of C4 photosynthesis: more than decarboxylating enzymes. Frontiers in Plant Science, 10: 247, doi: 10.3389/fpls.2019.00247.
|
|
|
| [58] |
Sage R F, Kubien D S. 2007. The temperature response of C(3) and C(4) photosynthesis Plant, Cell & Environment, 30(9): 1086-1106.
doi: 10.1111/pce.2007.30.issue-9
|
|
|
| [59] |
Samiei L, Pahnehkolayi M D, Karimian Z, et al. 2020. Morpho-physiological responses of halophyte Climacoptera crassa to salinity and heavy metal stresses in in vitro condition. South African Journal of Botany, 131: 468-474.
doi: 10.1016/j.sajb.2020.03.037
|
|
|
| [60] |
Selinski J, Scheibe R. 2019. Malate valves: old shuttles with new perspectives. Plant Biology, 21(Suppl 1): 21-30.
doi: 10.1111/plb.2019.21.issue-S1
|
|
|
| [61] |
Shen L L, Tang K L, Wang W D, et al. 2022. Architecture of the chloroplast PSI-NDH supercomplex in Hordeum vulgare. Nature, 601: 649-654.
doi: 10.1038/s41586-021-04277-6
|
|
|
| [62] |
Shikanai T. 2016. Chloroplast NDH: a different enzyme with a structure similar to that of respiratory NADH dehydrogenase. Biochimica et Biophysica Acta (BBA) - Bioenergetics, 1857(7): 1015-1022.
doi: 10.1016/j.bbabio.2015.10.013
|
|
|
| [63] |
Shiri M, Rabhi M, El Amrani A, et al. 2015. Cross-tolerance to abiotic stresses in halophytes: application for phytoremediation of organic pollutants. Acta Physiologiae Plantarum, 37(10): 209, doi: 10.1007/s11738-015-1954-0.
|
|
|
| [64] |
Shiri M, Rabhi M, Abdelly C, et al. 2016. Moderate salinity reduced phenanthrene-induced stress in the halophyte plant model Thellungiella salsuginea compared to its glycophyte relative Arabidopsis thaliana: Cross talk and metabolite profiling. Chemosphere, 155: 453-462.
doi: S0045-6535(16)30566-5
pmid: 27139124
|
|
|
| [65] |
Singh J, Garai S, Das S, et al. 2022. Role of C4 photosynthetic enzyme isoforms in C3 plants and their potential applications in improving agronomic traits in crops. Photosynthesis Research, 154(3): 233-258.
doi: 10.1007/s11120-022-00978-9
|
|
|
| [66] |
Takabayashi A, Kishine M, Asada K, et al. 2005. Differential use of two cyclic electron flows around photosystem I for driving CO2-concentration mechanism in C4 photosynthesis. Proceedings of the National Academy of Sciences of the United States of America, 102(46): 16898-16903.
|
|
|
| [67] |
Tian W, Su C F, Zhang N, et al. 2024. Simulation of the physiological and photosynthetic characteristics of C3 and C4 plants under elevated temperature and CO2 concentration. Ecological Modelling, 495: 110805, doi: 10.1016/j.ecolmodel.2024.110805.
|
|
|
| [68] |
Tiwari Y K, Yadav S K. 2019. High temperature stress tolerance in maize (Zea mays L.): Physiological and molecular mechanisms. Journal of Plant Biology, 62: 93-102.
doi: 10.1007/s12374-018-0350-x
|
|
|
| [69] |
Toderich K N, Shuyskaya E V, Ismail S, et al. 2009. Phytogenic resources of halophytes of Central Asia and their role for rehabilitation of sandy desert degraded rangelands. Land Degradation & Development, 20(4): 386-396.
doi: 10.1002/ldr.v20:4
|
|
|
| [70] |
Tomar R S, Kataria S, Jajoo A. 2021. Behind the scene: Critical role of reactive oxygen species and reactive nitrogen species in salt stress tolerance. Journal of Agronomy and Crop Science, 207(4): 577-588.
doi: 10.1111/jac.v207.4
|
|
|
| [71] |
Tsuchida H, Tamai T, Fukayama H, et al. 2001. High level expression of C4-specific NADP-malic enzyme in leaves and impairment of photoautotrophic growth in a C3 plant, rice. Plant and Cell Physiology, 42(2): 138-145.
pmid: 11230567
|
|
|
| [72] |
Voon C P, Law Y S, Guan X Q, et al. 2021. Modulating the activities of chloroplasts and mitochondria promotes adenosine triphosphate production and plant growth. Quantitative Plant Biology, 2: e7, doi: 10.1017/qpb.2021.7.
|
|
|
| [73] |
Wang L, Wei J, Shi X Y, et al. 2023. Identification of the light-harvesting chlorophyll a/b binding protein gene family in peach (Prunus persica L.) and their expression under drought stress. Genes, 14(7): 1475, doi: 10.3390/genes14071475.
|
|
|
| [74] |
Wang Y, Bräutigam A, Weber A P M, et al. 2014. Three distinct biochemical subtypes of C4 photosynthesis? A modelling analysis. Journal of Experimental Botany, 65(13): 3567-3578.
doi: 10.1093/jxb/eru058
pmid: 24609651
|
|
|
| [75] |
Wen Z B, Zhang M L. 2017. Possible involvement of phosphoenolpyruvate carboxylase and NAD-malic enzyme in response to drought stress. A case study: a succulent nature of the C4-NAD-ME type desert plant, Salsola lanata (Chenopodiaceae). Functional Plant Biology, 44(12): 1219-1228.
|
|
|
| [76] |
Yan K, Chen P, Shao H, et al. 2012. Responses of photosynthesis and photosystem II to higher temperature and salt stress in sorghum. Journal of Agronomy and Crop Science, 198(3): 218-225.
doi: 10.1111/jac.2012.198.issue-3
|
|
|
| [77] |
Yıldız M, Terzi H. 2021. Comparative analysis of salt-induced changes in the root physiology and proteome of the xero-halophyte Salsola crassa. Brazilian Journal of Botany, 44: 33-42.
doi: 10.1007/s40415-020-00695-4
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|