| Research article |
|


|
|
|
| Potential distribution of Haloxylon ammodendron in Central Asia under climate change |
CHEN Zhuo1,2, SHAO Minghao2,3, HU Zihao4, GAO Xin1,2,*( ), LEI Jiaqiang1,2 ), LEI Jiaqiang1,2 |
1Key Laboratory of Ecological Safety and Sustainable Development in Arid Lands, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China
2University of Chinese Academy of Sciences, Beijing 100049, China
3National Engineering Technology Research Center for Desert-Oasis Ecological Construction, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China
4University of Emergency Management, Beijing 101601, China |
|
|
|
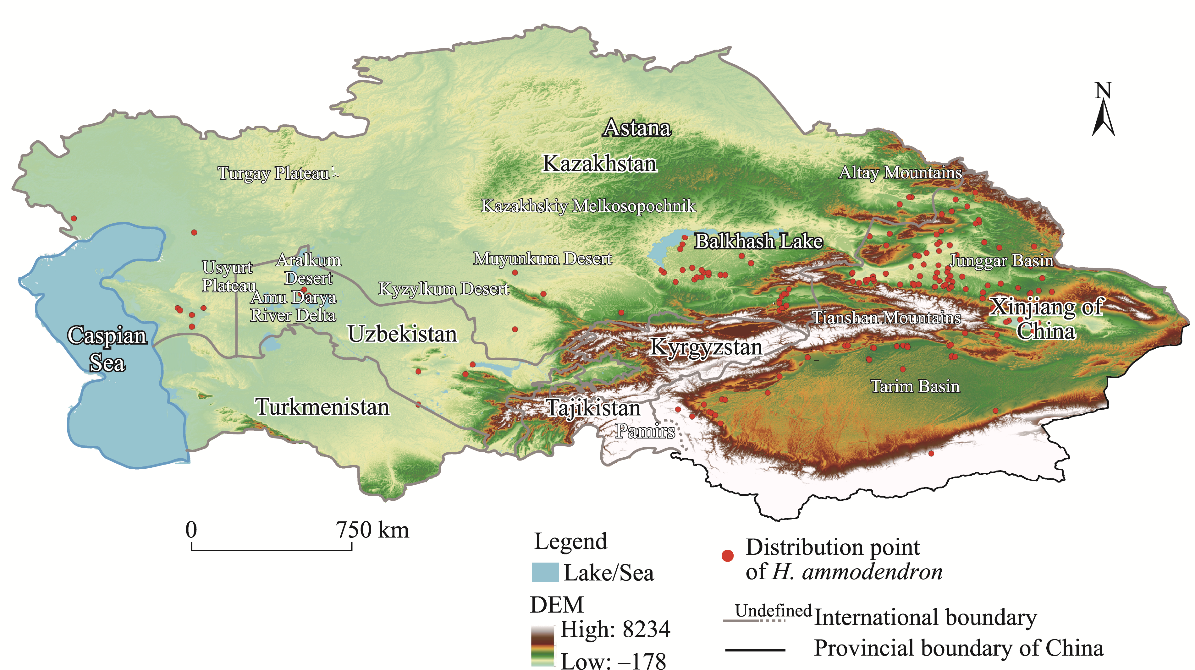
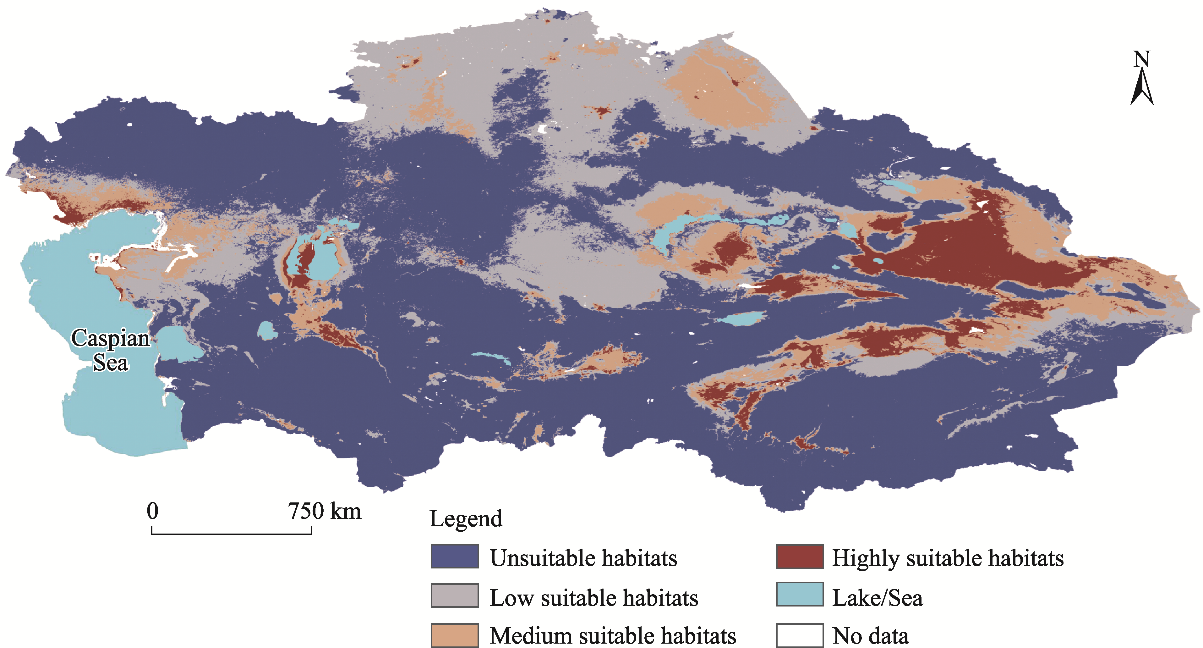
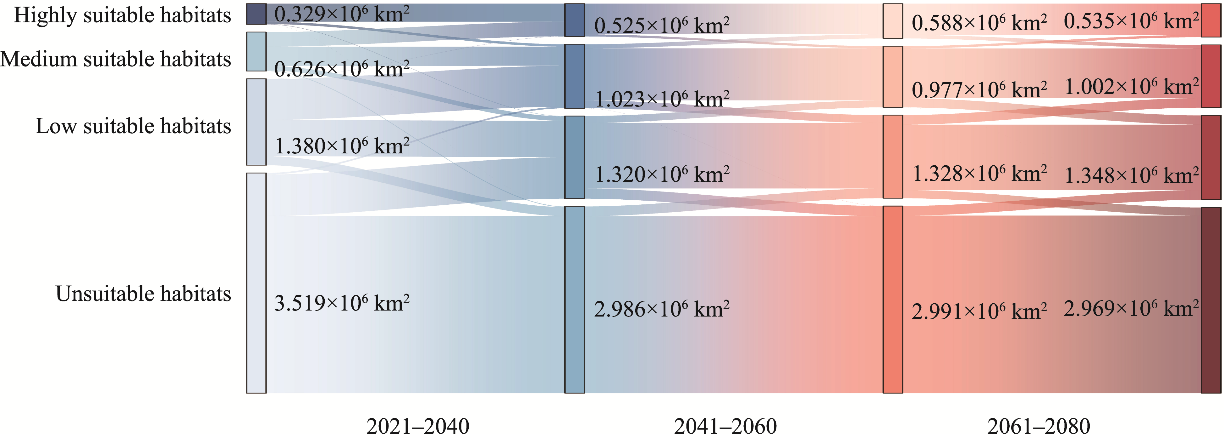
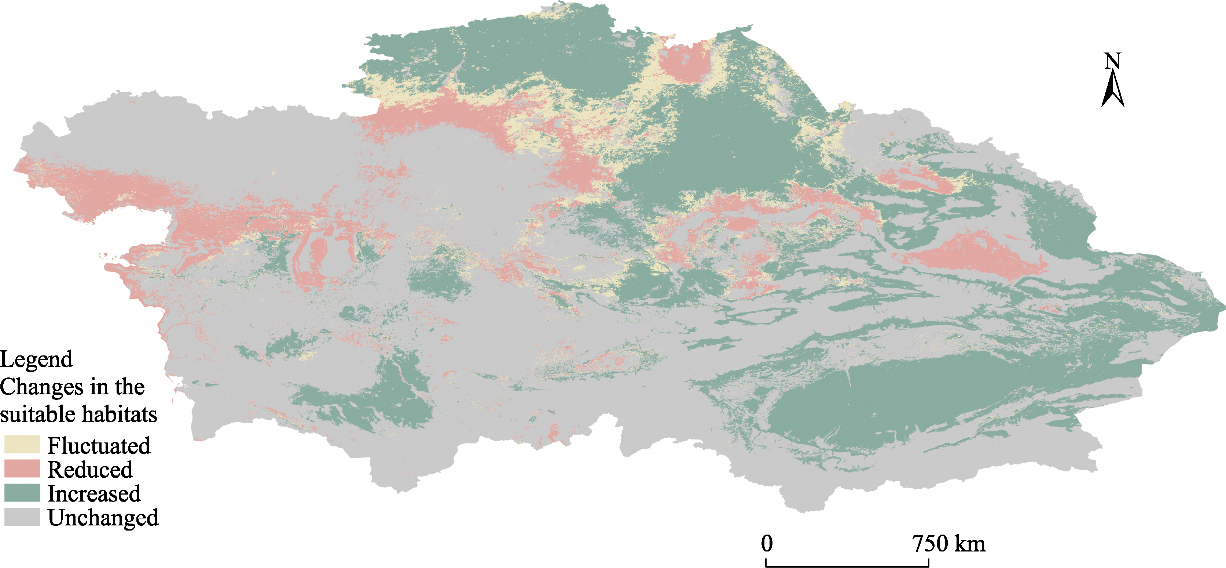
Abstract Understanding the spatial distribution of plant species and their dynamic changes in arid areas is crucial for addressing the challenges posed by climate change. Haloxylon ammodendron shelterbelts are essential for the protection of plant resources and the control of desertification in Central Asia. Thus far, the potential suitable habitats of H. ammodendron in Central Asia are still uncertain in the future under global climate change conditions. This study utilised the maximum entropy (MaxEnt) model to combine the current distribution data of H. ammodendron with its growth-related data to analyze the potential distribution pattern of H. ammodendron across Central Asia. The results show that there are suitable habitats of H. ammodendron in the Aralkum Desert, northern slopes of the Tianshan Mountains, and the upstream of the Tarim River and western edge of the Taklimakan Desert in the Tarim Basin under the current climate conditions. The period from 2021 to 2040 is projected to undergo significant changes in the suitable habitat area of H. ammodendron in Central Asia, with a projected 15.0% decrease in the unsuitable habitat area. Inland areas farther from the ocean, such as the Caspian Sea and Aralkum Desert, will continue to experience a decrease in the suitable habitats of H. ammodendron. Regions exhibiting frequent fluctuations in the habitat suitability levels are primarily found along the axis stretching from Astana to Kazakhskiy Melkosopochnik in Kazakhstan. These regions can transition into suitable habitats under varying climate conditions, requiring the implementation of appropriate human intervention measures to prevent desertification. Future climate conditions are expected to cause an eastward shift in the geometric centre of the potential suitable habitats of H. ammodendron, with the extent of this shift amplifying alongside more greenhouse gas emissions. This study can provide theoretical support for the spatial configuration of H. ammodendron shelterbelts and desertification control in Central Asia, emphasising the importance of proactive measures to adapt to climate change in the future.
|
|
Received: 01 April 2024
Published: 30 September 2024
|
|
|
|
Corresponding Authors:
*GAO Xin (E-mail: gaoxin@ms.xjb.ac.cn)
|
|
|
| [1] |
Abdi E, Saleh H R, Majnonian B, et al. 2019. Soil fixation and erosion control by Haloxylon persicum roots in arid lands, Iran. Journal of Arid Land, 11(1): 86-96.
|
|
|
| [2] |
Ahmadi M, Hemami M R, Kaboli M, et al. 2023. MaxEnt brings comparable results when the input data are being completed: Model parameterization of four species distribution models. Ecology and Evolution, 13(2): e9827, doi: 10.1002/ece3.9827.
|
|
|
| [3] |
Bohovic R, Dobrovolny P, Klein D. 2016. The spatial and temporal dynamics of remotely-sensed vegetation phenology in Central Asia in the 1982-2011 period. European Journal of Remote Sensing, 49(1): 279-299.
|
|
|
| [4] |
Bouchet P J, Miller D L, Roberts J J, et al. 2020. Dsmextra: Extrapolation assessment tools for density surface models. Methods in Ecology and Evolution, 11(11): 1464-1469.
|
|
|
| [5] |
Chen Z, Gao X, Lei J Q. 2022. Dust emission and transport in the Aral Sea region. Geoderma, 428: 116177, doi: 10.1016/j.geoderma.2022.116177.
|
|
|
| [6] |
Davi N K, D'Arrigo R, Jacoby G C, et al. 2015. A long-term context (931-2005 C.E.) for rapid warming over Central Asia. Quaternary Science Reviews, 121: 89-97.
|
|
|
| [7] |
Dawson T P, Jackson S T, House J I, et al. 2011. Beyond predictions: biodiversity conservation in a changing climate. Science, 332(6025): 53-58.
doi: 10.1126/science.1200303
pmid: 21454781
|
|
|
| [8] |
de Beurs K M, Henebry G M, Owsley B C, et al. 2015. Using multiple remote sensing perspectives to identify and attribute land surface dynamics in Central Asia 2001-2013. Remote Sensing of Environment, 170: 48-61.
|
|
|
| [9] |
De Marco Júnior P, Nóbrega C C. 2018. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE, 13(9): e0202403, doi: 10.1371/journal.pone.0202403.
|
|
|
| [10] |
Dillon M E, Wang G, Huey R B. 2010. Global metabolic impacts of recent climate warming. Nature, 467(7316): 704-706.
|
|
|
| [11] |
Etherington T R. 2019. Mahalanobis distances and ecological niche modelling: correcting a chi-squared probability error. PeerJ, 7: e6678, doi: 10.7717/peerj.6678.
|
|
|
| [12] |
Fang J Q, Shi J F, Zhang P, et al. 2024. Potential distribution projections for Senegalia senegal (L.) Britton under climate change scenarios. Forests, 15(2): 379, doi: 10.3390/f15020379.
|
|
|
| [13] |
Guillera-Arroita G, Lahoz-Monfort J, Elith J. 2014. MaxEnt is not a presence-absence method: a comment on Thibaud et al. Methods in Ecology and Evolution, 5(11): 1192-1197.
|
|
|
| [14] |
Guisan A, Edwards Jr T C, Hastie T. 2002. Generalized linear and generalized additive models in studies of species distributions: setting the scene. Ecological Modelling, 157(2-3): 89-100.
|
|
|
| [15] |
Hirzel A H, Hausser J, Chessel D, et al. 2002. Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecology, 83(7): 2027-2036.
|
|
|
| [16] |
Hu Z Y, Zhang C, Hu Q, et al. 2014. Temperature changes in Central Asia from 1979 to 2011 based on multiple datasets. Journal of Climate, 27(3): 1143-1167.
|
|
|
| [17] |
Intergovernmental Panel on Climate Change (IPCC). 2023. Climate Change 2023:Synthesi Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva: IPCC, 35-115.
|
|
|
| [18] |
Jiang L L, Jiapaer G, Bao A M, et al. 2017. Vegetation dynamics and responses to climate change and human activities in Central Asia. Science of the Total Environment, 599-600: 967-980.
|
|
|
| [19] |
Kang J F, Liu M S, Qu M K, et al. 2023. Identifying the potential soil pollution areas derived from the metal mining industry in China using MaxEnt with mine reserve scales (MaxEnt_MRS). Environmental Pollution, 329: 121687, doi: 10.1016/j.envpol.2023.121687.
|
|
|
| [20] |
Leroy B, Delsol R, Hugueny B, et al. 2018. Without quality presence-absence data, discrimination metrics such as TSS can be misleading measures of model performance. Journal of Biogeography, 45(9): 1994-2002.
|
|
|
| [21] |
Li C J, Han H, Ablimiti M, et al. 2022. Morphological and physiological responses of desert plants to drought stress in a man-made landscape of the Taklimakan Desert shelter belt. Ecological Indicators, 140: 109037, doi: 10.1016/j.ecolind.2022.109037.
|
|
|
| [22] |
Li J Y, Chang H, Liu T, et al. 2019. The potential geographical distribution of Haloxylon across Central Asia under climate change in the 21st century. Agricultural and Forest Meteorology, 275: 243-254.
|
|
|
| [23] |
Lioubimtseva E. 2015. A multi-scale assessment of human vulnerability to climate change in the Aral Sea basin. Environmental Earth Sciences, 73: 719-729.
|
|
|
| [24] |
Mannocci L, Roberts J J, Miller D L, et al. 2017. Extrapolating cetacean densities to quantitatively assess human impacts on populations in the high seas. Conservation Biology, 31(3): 601-614.
doi: 10.1111/cobi.12856
pmid: 27775847
|
|
|
| [25] |
Mannocci L, Roberts J J, Halpin P N, et al. 2018. Assessing cetacean surveys throughout the Mediterranean Sea: A gap analysis in environmental space. Scientific Reports, 8(1): 3126, doi: 10.1038/s41598-018-19842-9.
|
|
|
| [26] |
Marmion M, Parviainen M, Luoto M, et al. 2009. Evaluation of consensus methods in predictive species distribution modelling. Diversity and Distributions, 15(1): 59-69.
|
|
|
| [27] |
Mi C R, Huettmann F, Guo Y M, et al. 2017. Why choose Random Forest to predict rare species distribution with few samples in large undersampled areas? Three Asian crane species models provide supporting evidence. PeerJ, 5: e2849, doi: 10.7287/peerj.preprints.2517v1.
|
|
|
| [28] |
Micklin P. 2007. The Aral Sea disaster. Annual Review of Earth and Planetary Sciences, 35: 47-72.
|
|
|
| [29] |
Miller D L, Burt M L, Rexstad E A, et al. 2013. Spatial models for distance sampling data: recent developments and future directions. Methods in Ecology and Evolution, 4(11): 1001-1010.
|
|
|
| [30] |
Mohammat A, Wang X H, Xu X T, et al. 2013. Drought and spring cooling induced recent decrease in vegetation growth in Inner Asia. Agricultural and Forest Meteorology, 178-179: 21-30.
|
|
|
| [31] |
Muscarella R, Galante P J, Soley-Guardia M, et al. 2015. Enmeval: an R package for conducting spatially independent evaluations and estimating optimal model complexity for maxent ecological niche models. Methods in Ecology and Evolution, 5(11): 1198-1205.
|
|
|
| [32] |
Paolo D, Bhattachan A, Kyle F, et al. 2013. Global desertification: Drivers and feedbacks. Advances in Water Resources, 51: 326-344.
|
|
|
| [33] |
Pecchi M, Marchi M, Moriondo M, et al. 2020. Potential impact of climate change on the forest coverage and the spatial distribution of 19 key forest tree species in Italy under RCP4.5 IPCC trajectory for 2050s. Forests, 11(9): 934, doi: 10.3390/f11090934.
|
|
|
| [34] |
Phillips S J, Anderson R P, Schapire R E. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190(3-4): 231-259.
|
|
|
| [35] |
Qi K, Zhu J J, Zheng X, et al. 2023. Impacts of the world's largest afforestation program (Three-North Afforestation Program) on desertification control in sandy land of China. GIScience & Remote Sensing, 60(1): 2167574, doi: 10.1080/15481603.2023.2167574.
|
|
|
| [36] |
Rengasamy P. 2006. World salinization with emphasis on Australia. Journal of Experimental Botany, 57(5): 1017-1023.
doi: 10.1093/jxb/erj108
pmid: 16510516
|
|
|
| [37] |
Ruan X, Wang Q, Chen Y, et al. 2006. Physio-ecological response of Haloxylon persicum photosynthetic shoots to drought stress. Frontiers of Forestry in China, 1: 176-181.
|
|
|
| [38] |
Shao M H, Wang L, Li B W, et al. 2022. Maxent modeling for identifying the nature reserve of Cistanche deserticola Ma under effects of the host (Haloxylon Bunge) forest and climate changes in Xinjiang, China. Forests, 13(2): 189, doi: 10.3390/f13020189.
|
|
|
| [39] |
Shao Y. 2008. Physics and Modelling of Wind Erosion. Dordrecht: Springer Netherlands, 456, doi: 10.1007/978-1-4020-8895-7.
|
|
|
| [40] |
Sillero N, Gonçalves-Seco L. 2014. Spatial structure analysis of a reptile community with airborne LiDAR data. International Journal of Geographical Information Science, 28(8): 1709-1722.
|
|
|
| [41] |
Sofaer H, Jarnevich C, Pearse I S, et al. 2019. Development and Delivery of Species Distribution Models to Inform Decision-Making. BioScience, 69(7): 544-557.
doi: 10.1093/biosci/biz045
|
|
|
| [42] |
Song Y L, Zeng K W, Jiang Y, et al. 2021. Cistanches Herba, from an endangered species to a big brand of Chinese medicine. Medicinal Research Reviews, 41(3): 1539-1577.
doi: 10.1002/med.21768
pmid: 33521978
|
|
|
| [43] |
Sorichetta A, Hornby G M, Stevens F R, et al. 2015. High-resolution gridded population datasets for Latin America and the Caribbean in 2010, 2015, and 2020. Scientific Data, 2: 150045, doi: 10.1038/sdata.2015.45.
|
|
|
| [44] |
Sun J J, Qiu H J, Guo J H, et al. 2020. Modeling the potential distribution of Zelkova schneideriana under different human activity intensities and climate change patterns in China. Global Ecology and Conservation, 21: e00840, doi: 10.1016/j.gecco.2019.e00840.
|
|
|
| [45] |
Tao Y, Wu G L, Zhang Y M. 2017. Dune-scale distribution pattern of herbaceous plants and their relationship with environmental factors in a saline-alkali desert in Central Asia. Science of the Total Environment, 576: 473-480.
|
|
|
| [46] |
Tarhouni M, Salem F B, Belgacem A O, et al. 2010. Acceptability of plant species along grazing gradients around watering points in Tunisian arid zone. Flora-Morphology, Distribution, Functional Ecology of Plants, 205(7): 454-461.
|
|
|
| [47] |
Tomislav H. 2018. Sand content in % (kg/kg) at 6 standard depths (0, 10, 30, 60, 100 and 200 cm) at 250 m resolution (v0.2)[Data set]. Zenodo. https://doi.org/10.5281/zenodo.2525662.
|
|
|
| [48] |
Tomislav H, Surya G. 2019. Soil water content (volumetric %) for 33 kPa and 1500 kPa suctions predicted at 6 standard depths (0, 10, 30, 60, 100 and 200 cm) at 250 m resolution (v0.1)[Data set]. Zenodo. https://doi.org/10.5281/zenodo.2784001.
|
|
|
| [49] |
Trampetti F, Pereira C, Rodrigues M J, et al. 2019. Exploring the halophyte Cistanche phelypaea (L.) Cout as a source of health promoting products: In vitro antioxidant and enzyme inhibitory properties, metabolomic profile and computational studies. Journal of Pharmaceutical and Biomedical Analysis, 165: 119-128.
|
|
|
| [50] |
Velasco J A, González-Salazar C. 2019. Akaike information criterion should not be a ''test'' of geographical prediction accuracy in ecological niche modelling. Ecological Informatics, 51: 25-32.
doi: 10.1016/j.ecoinf.2019.02.005
|
|
|
| [51] |
Wang L J, Zhao C Y, Li J, et al. 2009. Study on species diversity in early-spring in Haloxylon desert in north Fukang region, Xinjiang. Arid Zone Research, 26(4): 574-581. (in Chinese)
|
|
|
| [52] |
Xiao F J, Liu Q F, Qin Y. 2024. Predicting the potential distribution of Haloxylon ammodendron under climate change scenarios using machine learning of a maximum entropy model. Biology, 13(1): 3, doi: 10.3390/biology13010003.
|
|
|
| [53] |
Yang W J, Sun S X, Wang N X, et al. 2023. Dynamics of the distribution of invasive alien plants (Asteraceae) in China under climate change. Science of the Total Environment, 903: 166260, doi: 10.1016/j.scitotenv.2023.166260.
|
|
|
| [54] |
Zhang X X, Claiborn C, Lei J Q, et al. 2020. Aeolian dust in Central Asia: Spatial distribution and temporal variability. Atmospheric Environment, 238: 117734, doi: 10.1016/j.atmosenv.2020.117734.
|
|
|
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|